Nature of a Nerve Impulse
Nature of a Nerve Impulse
A nerve impulse is an electro-chemical message of neurons, the common functional denominator of all nervous system activity. Despite the incredible complexity of the nervous system of many animals, nerve impulses are basically alike in all neurons and in all animals. An impulse is an “all-ornone” phenomenon; either the fiber is conducting an impulse, or it is not. Because all impulses are alike, the only way a nerve fiber can vary its signal is by changing the frequency of impulse conduction. Frequency change is the language of a nerve fiber. A fiber may conduct no impulses at all or very few per second up to a maximum approaching 1000 per second. The higher the frequency (or rate) of conduction, the greater is the level of excitation.
Resting Membrane Potential
Membranes of neurons, like all cellular membranes, have a special permeability that creates ionic imbalances. The interstitial fluid surrounding neurons contains relatively high concentrations of sodium (Na+) and chloride (Cl−) ions, but a low concentration of potassium ions (K+) and large impermeable anions with negative charge. Inside the neuron, the ratio is reversed: the K+ and impermeable anion concentration is high, but the Na+ and Cl− concentrations are low (Figure 35-4; see also Figure 33-1B, ) These differences are pronounced; there is approximately 10 times more Na+ outside than in and 25 to 30 times more K+ inside than out.
When at rest, the membrane of a neuron is selectively permeable to K+, which can traverse the membrane through special potassium channels. The permeability to Na+ is nearly zero because the Na+ channels are closed in a resting membrane. Potassium ions tend to diffuse outward through the membrane, following the gradient of potassium concentration. Very quickly the positive charge outside reaches a level that prevents any more K+ from diffusing out of the axon (because like charges repel each other), and because the large anions cannot pass through the membrane, the positively charged potassium ions are drawn back into the cell. Now the resting membrane is at equilibrium, with an electrical gradient that exactly balances the concentration gradient. This resting membrane potential is usually −70 mV (millivolts), with the inside of the membrane negative with respect to the outside.
Action Potential
A nerve impulse is a rapidly moving change in electrical membrane potential called an action potential (Figure 35-5). It is a very rapid and brief depolarization of the membrane of the nerve fiber. In most nerve fibers, the action potential does not simply return the membrane potential to zero, but instead overshoots zero. In other words, the membrane potential reverses for an instant so that the outside becomes negative compared with the inside. Then, as the action potential moves ahead, the membrane returns to its normal resting membrane potential, ready to conduct another - impulse. The entire event occupies approximately a millisecond. Perhaps the most significant property of the nerve impulse is that it is selfpropagating; once started the impulse moves ahead automatically, much like the burning of a fuse.
What causes the reversal of polarity in the cell membrane during passage of an action potential? We have seen that the resting membrane potential depends on the high membrane permeability (leakiness) to K+, some 50 to 70 times greater than the permeability to Na+. When the action potential arrives at a given point, Na+ channels suddenly open, permitting a flood of Na+ to diffuse into the axon from the outside, moving down the concentration gradient for Na+. Actually only a very minute amount of Na+ moves across the membrane— less than one-millionth of the Na+outside—but this sudden rush of positive ions cancels the local resting membrane potential. The membrane is depolarized, creating a minute electrical “hole.” Potassium ions, finding their electrical barrier gone, begin to move outside the cell. Then, as the action potential passes, the membrane quickly regains its resting properties. It becomes once again practically impermeable to Na+ and the outward movement of K+ is checked. Thus, the rising phase of the action potential is associated with rapid influx (inward movement) of Na+(Figure 35-5). When the action potential reaches its peak, Na+ permeability is restored to normal, and K+ permeability briefly increases above the resting level. Increased potassium permeability causes the action potential to drop rapidly toward the resting membrane level, during the repolarization phase. The membrane is now ready to transmit another nerve impulse.
Sodium Pump
A resting cell membrane has a very low permeability to Na+. Nevertheless some Na+ leaks through it, even in the resting condition. When the axon is active, during an action potential, Na+ flows inward with each passing impulse. If not removed, the accumulation of Na+ inside the axon would cause the resting membrane potential of the fiber to decay. This decay is prevented by sodium pumps, each a complex of protein subunits embedded in the plasma membrane of the axon (see Figure 3-19,). Each sodium pump uses energy in ATP to transport sodium from the inside to the outside of the membrane. The sodium pump in nerve axons, as in many other cell membranes, also moves K+ into the axon while it is moving Na+ out. Thus, it is a sodium-potassium exchange pump that helps to restore the ion gradients of both Na+ and K+. The astrocytes (mentioned earlier) help to maintain the correct balance of ions surrounding neurons by sweeping away excess potassium produced during neuronal activity.
High-Speed Conduction
Although the ionic and electrical events associated with action potentials are much the same throughout the animal kingdom, conduction velocities vary enormously from nerve to nerve and from animal to animal—from as slow as 0.1 m/sec in sea anemones to as fast as 120 m/sec in some mammalian motor axons. The speed of conduction is closely related to the diameter of the axon. Small axons conduct slowly because internal resistance to current flow is high. In most invertebrates, where fast conduction velocities are important for quick response, such as in locomotion to capture prey or to avoid capture, axon diameters are larger. The giant axon of squids is nearly 1 mm in diameter and carries impulses 10 times faster than ordinary fibers in the same animal. A squid’s giant axon innervates the animal’s mantle musculature and is used for powerful mantle contractions when the animal swims by jet propulsion. Similar giant axons enable earthworms, which are normally slow-moving animals, to withdraw almost instantaneously into their burrows when startled.
Some invertebrates, including prawns and insects, also have fast fibers invested with multiple layers of a myelin-like substance that is interrupted at intervals much like myelinated fibers of vertebrates. Conduction rates, though not as fast as vertebrate saltatory conduction, are much faster than unmyelinated fibers of the same diameter in other invertebrates.
Vertebrates do not possess giant axons, but they can achieve highconduction velocities in another way, by a cooperative relationship between axons and the investing layers of myelin laid down by the Schwann cells or oligodendrocytes described earlier. Insulating myelin sheaths are interrupted at intervals by nodes (called nodes of Ranvier) where the surface of the axon is exposed to fluid surrounding the nerve. In these myelinated fibers the action potential depolarizes the axon membrane only at the nodes because the myelin sheath prevents depolarization elsewhere (Figure 35-6). The ion pumps and channels that move ions across the membrane are concentrated in each node. Once an action potential starts down an axon, depolarization of the first node initiates an electrical current that stretches out to the neighboring node, causing it to depolarize and trigger an action potential. Thus the action potential leaps from node to node, a kind of conduction called saltatory (L. salto, to dance, leap). The gain in efficiency as compared with nonmyelinated fibers is impressive. For example, a frog myelinated axon only 12 µm in diameter conducts nerve impulses at the same speed as a squid axon 350 µm in diameter.
A nerve impulse is an electro-chemical message of neurons, the common functional denominator of all nervous system activity. Despite the incredible complexity of the nervous system of many animals, nerve impulses are basically alike in all neurons and in all animals. An impulse is an “all-ornone” phenomenon; either the fiber is conducting an impulse, or it is not. Because all impulses are alike, the only way a nerve fiber can vary its signal is by changing the frequency of impulse conduction. Frequency change is the language of a nerve fiber. A fiber may conduct no impulses at all or very few per second up to a maximum approaching 1000 per second. The higher the frequency (or rate) of conduction, the greater is the level of excitation.
Resting Membrane Potential
Membranes of neurons, like all cellular membranes, have a special permeability that creates ionic imbalances. The interstitial fluid surrounding neurons contains relatively high concentrations of sodium (Na+) and chloride (Cl−) ions, but a low concentration of potassium ions (K+) and large impermeable anions with negative charge. Inside the neuron, the ratio is reversed: the K+ and impermeable anion concentration is high, but the Na+ and Cl− concentrations are low (Figure 35-4; see also Figure 33-1B, ) These differences are pronounced; there is approximately 10 times more Na+ outside than in and 25 to 30 times more K+ inside than out.
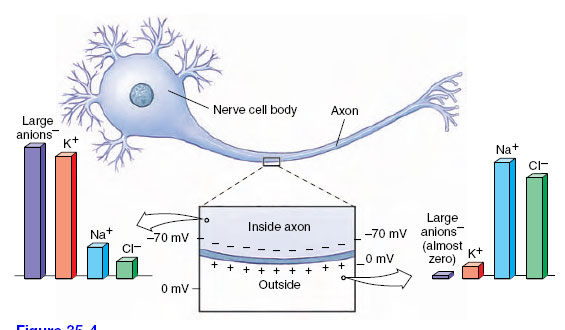 |
| Figure 35-4 Ionic composition inside and outside a resting nerve cell. An active sodium-potassium exchange pump located in the cell membrane drives sodium to the outside, keeping its concentration low inside. Potassium concentration is high inside. Although the membrane is “leaky” to potassium, this ion is held inside by the repelling positive charge outside the membrane, and its attraction to large negatively charged anions inside the membrane, which cannot leave the cell. |
When at rest, the membrane of a neuron is selectively permeable to K+, which can traverse the membrane through special potassium channels. The permeability to Na+ is nearly zero because the Na+ channels are closed in a resting membrane. Potassium ions tend to diffuse outward through the membrane, following the gradient of potassium concentration. Very quickly the positive charge outside reaches a level that prevents any more K+ from diffusing out of the axon (because like charges repel each other), and because the large anions cannot pass through the membrane, the positively charged potassium ions are drawn back into the cell. Now the resting membrane is at equilibrium, with an electrical gradient that exactly balances the concentration gradient. This resting membrane potential is usually −70 mV (millivolts), with the inside of the membrane negative with respect to the outside.
Action Potential
A nerve impulse is a rapidly moving change in electrical membrane potential called an action potential (Figure 35-5). It is a very rapid and brief depolarization of the membrane of the nerve fiber. In most nerve fibers, the action potential does not simply return the membrane potential to zero, but instead overshoots zero. In other words, the membrane potential reverses for an instant so that the outside becomes negative compared with the inside. Then, as the action potential moves ahead, the membrane returns to its normal resting membrane potential, ready to conduct another - impulse. The entire event occupies approximately a millisecond. Perhaps the most significant property of the nerve impulse is that it is selfpropagating; once started the impulse moves ahead automatically, much like the burning of a fuse.
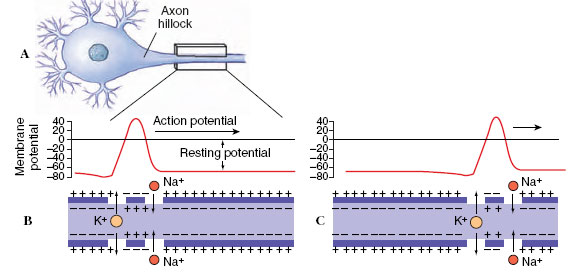 |
| Figure 35-5 Conduction of action potential of a nerve impulse. The action potential originates in the axon hillock of the neuron (A) and moves toward the right. B and C show the electrical event and associated changes in localized membrane permeability to sodium and potassium. The position of the action potential in C is shown about 4 milliseconds after B. When the impulse arrives at a point, sodium channels open, allowing sodium ions to enter. Sodium inflow reverses the membrane polarity, making the inner surface of the axon positive and the outside negative. Sodium channels then close and potassium channels open. Potassium ions can now restore the normal resting potential. |
What causes the reversal of polarity in the cell membrane during passage of an action potential? We have seen that the resting membrane potential depends on the high membrane permeability (leakiness) to K+, some 50 to 70 times greater than the permeability to Na+. When the action potential arrives at a given point, Na+ channels suddenly open, permitting a flood of Na+ to diffuse into the axon from the outside, moving down the concentration gradient for Na+. Actually only a very minute amount of Na+ moves across the membrane— less than one-millionth of the Na+outside—but this sudden rush of positive ions cancels the local resting membrane potential. The membrane is depolarized, creating a minute electrical “hole.” Potassium ions, finding their electrical barrier gone, begin to move outside the cell. Then, as the action potential passes, the membrane quickly regains its resting properties. It becomes once again practically impermeable to Na+ and the outward movement of K+ is checked. Thus, the rising phase of the action potential is associated with rapid influx (inward movement) of Na+(Figure 35-5). When the action potential reaches its peak, Na+ permeability is restored to normal, and K+ permeability briefly increases above the resting level. Increased potassium permeability causes the action potential to drop rapidly toward the resting membrane level, during the repolarization phase. The membrane is now ready to transmit another nerve impulse.
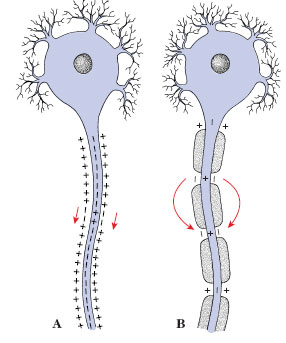 |
| Figure 35-6 Impulse conduction in unmyelinated and myelinated fibers. In unmyelinated fibers (A), the action potential spreads continuously along the entire length of the axon. In myelinated fibers (B) , the action potential leaps from node to node, bypassing the insulated portions of the fiber. This is saltatory conduction, which is much faster than continuous conduction. |
A resting cell membrane has a very low permeability to Na+. Nevertheless some Na+ leaks through it, even in the resting condition. When the axon is active, during an action potential, Na+ flows inward with each passing impulse. If not removed, the accumulation of Na+ inside the axon would cause the resting membrane potential of the fiber to decay. This decay is prevented by sodium pumps, each a complex of protein subunits embedded in the plasma membrane of the axon (see Figure 3-19,). Each sodium pump uses energy in ATP to transport sodium from the inside to the outside of the membrane. The sodium pump in nerve axons, as in many other cell membranes, also moves K+ into the axon while it is moving Na+ out. Thus, it is a sodium-potassium exchange pump that helps to restore the ion gradients of both Na+ and K+. The astrocytes (mentioned earlier) help to maintain the correct balance of ions surrounding neurons by sweeping away excess potassium produced during neuronal activity.
High-Speed Conduction
Although the ionic and electrical events associated with action potentials are much the same throughout the animal kingdom, conduction velocities vary enormously from nerve to nerve and from animal to animal—from as slow as 0.1 m/sec in sea anemones to as fast as 120 m/sec in some mammalian motor axons. The speed of conduction is closely related to the diameter of the axon. Small axons conduct slowly because internal resistance to current flow is high. In most invertebrates, where fast conduction velocities are important for quick response, such as in locomotion to capture prey or to avoid capture, axon diameters are larger. The giant axon of squids is nearly 1 mm in diameter and carries impulses 10 times faster than ordinary fibers in the same animal. A squid’s giant axon innervates the animal’s mantle musculature and is used for powerful mantle contractions when the animal swims by jet propulsion. Similar giant axons enable earthworms, which are normally slow-moving animals, to withdraw almost instantaneously into their burrows when startled.
Some invertebrates, including prawns and insects, also have fast fibers invested with multiple layers of a myelin-like substance that is interrupted at intervals much like myelinated fibers of vertebrates. Conduction rates, though not as fast as vertebrate saltatory conduction, are much faster than unmyelinated fibers of the same diameter in other invertebrates.
Vertebrates do not possess giant axons, but they can achieve highconduction velocities in another way, by a cooperative relationship between axons and the investing layers of myelin laid down by the Schwann cells or oligodendrocytes described earlier. Insulating myelin sheaths are interrupted at intervals by nodes (called nodes of Ranvier) where the surface of the axon is exposed to fluid surrounding the nerve. In these myelinated fibers the action potential depolarizes the axon membrane only at the nodes because the myelin sheath prevents depolarization elsewhere (Figure 35-6). The ion pumps and channels that move ions across the membrane are concentrated in each node. Once an action potential starts down an axon, depolarization of the first node initiates an electrical current that stretches out to the neighboring node, causing it to depolarize and trigger an action potential. Thus the action potential leaps from node to node, a kind of conduction called saltatory (L. salto, to dance, leap). The gain in efficiency as compared with nonmyelinated fibers is impressive. For example, a frog myelinated axon only 12 µm in diameter conducts nerve impulses at the same speed as a squid axon 350 µm in diameter.
Support our developers




