Photoreception: Vision
Photoreception: Vision
Light-sensitive receptors are called photoreceptors. These receptors range from simple light-sensitive cells scattered randomly on the body surface of many invertebrates (dermal light sense) to the exquisitely developed camera-type eye of vertebrates. Eyespots of astonishingly advanced organization appear even in some unicellular forms. That of the dinoflagellate Nematodinium bears a lens, a light-gathering chamber, and a photoreceptive pigment cup—all developed within a single-celled organism (Figure 35-30). The dermal light receptors of many invertebrates are of much simpler design. They are far less sensitive than optic receptors, but they are important in locomotory orientation, pigment distribution in chromatophores, photoperiodic adjustment of reproductive cycles, and other behavioral changes.
More highly organized eyes, many capable of excellent image formation, are based on one of two different principles: either a single-lens, camera-type eye such as those of cephalopod molluscs and vertebrates; or a multifaceted (compound) eye as in arthropods. Arthropod compound eyes are composed of many independent visual units called ommatidia (Figure 35-31). The eye of a bee contains about 15,000 of these units, each of which views a separate narrow sector of the visual field. Such eyes form a mosaic of images of varying brightness from the separate units. Resolution (the ability to see objects sharply) is poor compared with that of a vertebrate eye. A fruit fly, for example, must be closer than 3 cm to see another fruit fly as anything but a single spot. However, a compound eye is especially well suited to detecting motion, as anyone who has tried to swat a fly knows.
Eyes of certain annelids, molluscs, and all vertebrates are built like a camera—or rather we should say that a camera is modeled somewhat after vertebrate eyes. A camera-type eye contains in the front a light-tight chamber and lens system, which focuses an image of the visual field on a lightsensitive surface (the retina) in the back (Figure 35-32).
The spherical eyeball is built of three layers: (1) a tough outer white sclera for support and protection, (2) middle choroid coat, containing blood vessels for nourishment, and (3) light-sensitive retina (Figure 35-32). The cornea is a transparent anterior modification of the sclera. A circular, pigmented curtain, the iris, regulates the size of the light opening, the pupil. Just behind the iris is the lens, a transparent, elastic oval disc that, with the aid of ciliary muscles, can alter the curvature of the lens and bend light rays to focus an image on the retina. In terrestrial vertebrates the cornea actually does most of the bending of light rays, whereas the lens adjusts focus for near and far objects. Between cornea and lens is an outer chamber filled with watery aqueous humor; between lens and retina is a much larger inner chamber filled with viscous vitreous humor.
The retina is composed of several cell layers (Figure 35-33). The outermost layer, closest to the sclera, consists of pigment cells. Adjacent to this layer are the photoreceptors, rods and cones. Approximately 125 million rods and 1 million cones are present in each human eye. Cones are primarily concerned with color vision in ample light; rods, with colorless vision in dim light. Next is a network of intermediate neurons (bipolar, horizontal, and amacrine cells) that process and relay visual information from the photoreceptors to the ganglion cells whose axons form the optic nerve. The network permits much convergence, especially for rods. Information from several hundred rods may converge on a single ganglion cell, an adaptation that greatly increases the effectiveness of rods in dim light. Cones show very little convergence. By coordinating activities between different ganglion cells, and adjusting the sensitivities of bipolar cells, horizontal and amacrine cells improve overall contrast and quality of the visual image.
The fovea centralis or fovea, the region of keenest vision, is located in the center of the retina (Figure 35-32), in direct line with the center of the lens and cornea. It contains only cones, a vertebrate specialization for diurnal (daytime) vision. The acuity of an animal’s eyes depends on the density of cones in the fovea. The human fovea and that of a lion contain approximately 150,000 cones per square millimeter. But many water and field birds have up to 1 million cones per square millimeter. Their eyes are as good as our eyes would be if aided by eight-power binoculars.
One of several marvels of the vertebrate eye is its capacity to compress the enormous range of light intensities presented to it into a narrow range that can be handled by optic nerve fibers. Light intensity between a sunny noon and starlit night differs by more than 10 billion to 1. Rods quickly saturate with high light intensity, but cones do not; they shift their operating range with changing ambient light intensity so that a high-contrast image is perceived over a broad range of light conditions. This shift is made possible by complex interactions among the network of nerve cells that lie between the cones and the ganglion cells that generate the retinal output to the brain.
At the peripheral parts of the retina only rods are found. Rods are high-sensitivity receptors for dim light. At night, the cone-filled fovea is unresponsive to low levels of light and we become functionally color blind (“at night all cats are gray”). Under nocturnal conditions, the position of greatest visual acuity is not at the center of the fovea but at its edge. Thus it is easier to see a dim star at night by looking slightly to one side of it.
Light-sensitive receptors are called photoreceptors. These receptors range from simple light-sensitive cells scattered randomly on the body surface of many invertebrates (dermal light sense) to the exquisitely developed camera-type eye of vertebrates. Eyespots of astonishingly advanced organization appear even in some unicellular forms. That of the dinoflagellate Nematodinium bears a lens, a light-gathering chamber, and a photoreceptive pigment cup—all developed within a single-celled organism (Figure 35-30). The dermal light receptors of many invertebrates are of much simpler design. They are far less sensitive than optic receptors, but they are important in locomotory orientation, pigment distribution in chromatophores, photoperiodic adjustment of reproductive cycles, and other behavioral changes.
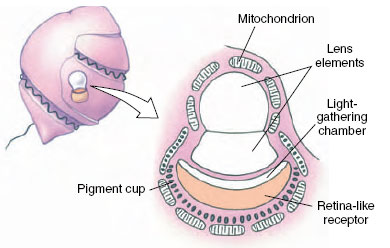 |
| Figure 35-30 Eyespot of the dinoflagellate Nematodinium. |
More highly organized eyes, many capable of excellent image formation, are based on one of two different principles: either a single-lens, camera-type eye such as those of cephalopod molluscs and vertebrates; or a multifaceted (compound) eye as in arthropods. Arthropod compound eyes are composed of many independent visual units called ommatidia (Figure 35-31). The eye of a bee contains about 15,000 of these units, each of which views a separate narrow sector of the visual field. Such eyes form a mosaic of images of varying brightness from the separate units. Resolution (the ability to see objects sharply) is poor compared with that of a vertebrate eye. A fruit fly, for example, must be closer than 3 cm to see another fruit fly as anything but a single spot. However, a compound eye is especially well suited to detecting motion, as anyone who has tried to swat a fly knows.
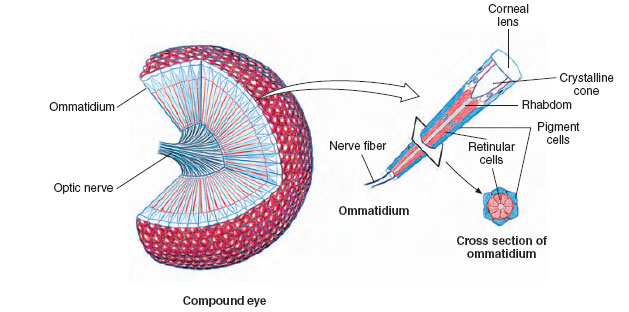 |
| Figure 35-31 Compound eye of an insect. A single ommatidium is shown enlarged at right |
Eyes of certain annelids, molluscs, and all vertebrates are built like a camera—or rather we should say that a camera is modeled somewhat after vertebrate eyes. A camera-type eye contains in the front a light-tight chamber and lens system, which focuses an image of the visual field on a lightsensitive surface (the retina) in the back (Figure 35-32).
The spherical eyeball is built of three layers: (1) a tough outer white sclera for support and protection, (2) middle choroid coat, containing blood vessels for nourishment, and (3) light-sensitive retina (Figure 35-32). The cornea is a transparent anterior modification of the sclera. A circular, pigmented curtain, the iris, regulates the size of the light opening, the pupil. Just behind the iris is the lens, a transparent, elastic oval disc that, with the aid of ciliary muscles, can alter the curvature of the lens and bend light rays to focus an image on the retina. In terrestrial vertebrates the cornea actually does most of the bending of light rays, whereas the lens adjusts focus for near and far objects. Between cornea and lens is an outer chamber filled with watery aqueous humor; between lens and retina is a much larger inner chamber filled with viscous vitreous humor.
The retina is composed of several cell layers (Figure 35-33). The outermost layer, closest to the sclera, consists of pigment cells. Adjacent to this layer are the photoreceptors, rods and cones. Approximately 125 million rods and 1 million cones are present in each human eye. Cones are primarily concerned with color vision in ample light; rods, with colorless vision in dim light. Next is a network of intermediate neurons (bipolar, horizontal, and amacrine cells) that process and relay visual information from the photoreceptors to the ganglion cells whose axons form the optic nerve. The network permits much convergence, especially for rods. Information from several hundred rods may converge on a single ganglion cell, an adaptation that greatly increases the effectiveness of rods in dim light. Cones show very little convergence. By coordinating activities between different ganglion cells, and adjusting the sensitivities of bipolar cells, horizontal and amacrine cells improve overall contrast and quality of the visual image.
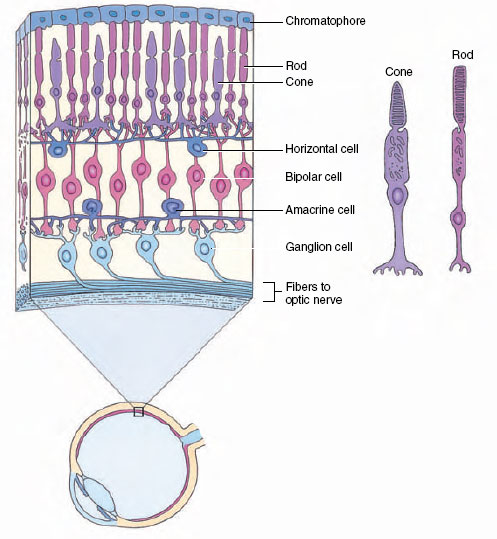 |
| Figure 35-33 Structure of a primate retina, showing organization of intermediate neurons that connect photoreceptor cells to ganglion cells of the optic nerve. |
The fovea centralis or fovea, the region of keenest vision, is located in the center of the retina (Figure 35-32), in direct line with the center of the lens and cornea. It contains only cones, a vertebrate specialization for diurnal (daytime) vision. The acuity of an animal’s eyes depends on the density of cones in the fovea. The human fovea and that of a lion contain approximately 150,000 cones per square millimeter. But many water and field birds have up to 1 million cones per square millimeter. Their eyes are as good as our eyes would be if aided by eight-power binoculars.
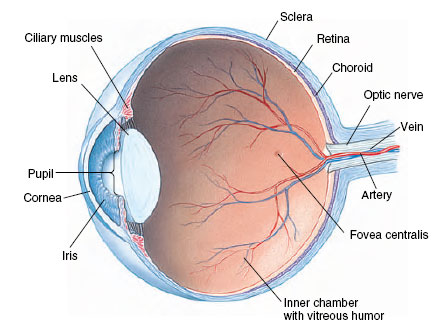 |
| Figure 35-32 Structure of the human eye. |
One of several marvels of the vertebrate eye is its capacity to compress the enormous range of light intensities presented to it into a narrow range that can be handled by optic nerve fibers. Light intensity between a sunny noon and starlit night differs by more than 10 billion to 1. Rods quickly saturate with high light intensity, but cones do not; they shift their operating range with changing ambient light intensity so that a high-contrast image is perceived over a broad range of light conditions. This shift is made possible by complex interactions among the network of nerve cells that lie between the cones and the ganglion cells that generate the retinal output to the brain.
At the peripheral parts of the retina only rods are found. Rods are high-sensitivity receptors for dim light. At night, the cone-filled fovea is unresponsive to low levels of light and we become functionally color blind (“at night all cats are gray”). Under nocturnal conditions, the position of greatest visual acuity is not at the center of the fovea but at its edge. Thus it is easier to see a dim star at night by looking slightly to one side of it.
Support our developers




