Synapses: Junctions Between Nerves
Synapses: Junctions
Between Nerves
When an action potential passes down an axon to its terminal, it must cross a small gap, the synapse (Gr. synapsis, contact, union), separating it from another neuron or an effector organ. Two distinct kinds of synapses are known: electrical and chemical.
Electrical synapses, although much less common than chemical synapses, have been demonstrated in both invertebrate and vertebrate groups. Electrical synapses are points at which ionic currents flow directly across a narrow gap junction (see Figure 3-15,) from one neuron to another. Electrical synapses show no time lag and consequently are important for escape reactions. They also have been observed in other excitable cell types, and form an important method of communication between cardiac muscle cells of the heart and smooth muscle cells (for example, the uterus,).
Much more complex than electrical synapses are chemical synapses, which contain packets of specialized chemicals called neurotransmitters. Neurons bringing impulses toward chemical synapses are called presynaptic neurons; those carrying impulses away are postsynaptic neurons. At a synapse, membranes are separated by a narrow gap, the synaptic cleft, having a width of approximately 20 nm.
The axon of most neurons divides at its end into many branches, each of which bears a synaptic knob that sits on the dendrites or cell body of the next neuron (Figure 35-7A). Because a single impulse coming down a nerve axon is transmitted along these many branches and synaptic endings on the next neuron, many impulses converge on the cell body at one instant. In addition, the axon terminations of many neurons may almost cover a nerve cell body and its dendrites with thousands of synapses. The 20 nm fluid-filled gap between presynaptic and postsynaptic membranes prevents action potentials from spreading directly to the postsynaptic neuron. Instead the synaptic knobs secrete a specific neurotransmitter that communicates chemically with the postsynaptic cell. One of the most common neurotransmitters of the peripheral nervous system is acetylcholine, which illustrates typical synaptic transmission. Inside the synaptic knobs are numerous tiny synaptic vesicles, each containing several thousand molecules of acetylcholine. Evidence suggests that when an impulse arrives at a terminal knob a sequence of events occurs as portrayed in Figure 35-8. The action potential causes an inward movement of calcium (Ca+) ions through channels in the synaptic knob membrane and this induces exocytosis of some neurotransmitter-filled synaptic vesicles. Acetylcholine molecules diffuse across the gap in a fraction of a millisecond and bind briefly to receptor molecules on ion channels in the postsynaptic membrane. This creates a voltage change in the postsynaptic membrane. Whether the voltage change is large enough to trigger a postsynaptic potential depends on how many acetylcholine molecules are released and how many channels are opened. Acetylcholine is rapidly destroyed by the enzyme acetylcholinesterase, which converts acetylcholine into acetate and choline. If not inactivated in this way, the neurotransmitter would continue to stimulate indefinitely. Organophosphate insecticides (such as malathion) and certain military nerve gases are poisonous for precisely this reason; they block acetylcholinesterase. The final step in the sequence is reabsorption of choline into the presynaptic terminal, resynthesis of acetylcholine and its storage in synaptic vesicles, ready to respond to another impulse.
Many different chemical neurotransmitters have been identified in both vertebrate and invertebrate nervous systems. Some, such as acetylcholine, norepinephrine, and glutamate, depolarize postsynaptic membranes; they are released at excitatory synapses. Other neurotransmitters, such as gamma aminobutyric acid (GABA), hyperpolarize postsynaptic membranes; thereby stabilizing them against depolarization. These neurotransmitters are released at inhibitory synapses. Neurons in the central nervous system have both excitatory and inhibitory synapses among the hundreds or thousands of synaptic knobs on the dendrites and cell body of each neuron.
The net balance of all excitatory and inhibitory inputs received by a postsynaptic cell determines whether it generates an action potential (Figure 35-8). If many excitatory impulses are received at one time, they can reduce the resting membrane potential enough in the postsynaptic membrane to elicit an action potential. Inhibitory impulses, however, stabilize the postsynaptic membrane, making it less likely that an action potential will be generated. The synapse is a crucial part of the decision-making equipment of the central nervous system, modulating flow of information from one neuron to the next.
When an action potential passes down an axon to its terminal, it must cross a small gap, the synapse (Gr. synapsis, contact, union), separating it from another neuron or an effector organ. Two distinct kinds of synapses are known: electrical and chemical.
Electrical synapses, although much less common than chemical synapses, have been demonstrated in both invertebrate and vertebrate groups. Electrical synapses are points at which ionic currents flow directly across a narrow gap junction (see Figure 3-15,) from one neuron to another. Electrical synapses show no time lag and consequently are important for escape reactions. They also have been observed in other excitable cell types, and form an important method of communication between cardiac muscle cells of the heart and smooth muscle cells (for example, the uterus,).
Much more complex than electrical synapses are chemical synapses, which contain packets of specialized chemicals called neurotransmitters. Neurons bringing impulses toward chemical synapses are called presynaptic neurons; those carrying impulses away are postsynaptic neurons. At a synapse, membranes are separated by a narrow gap, the synaptic cleft, having a width of approximately 20 nm.
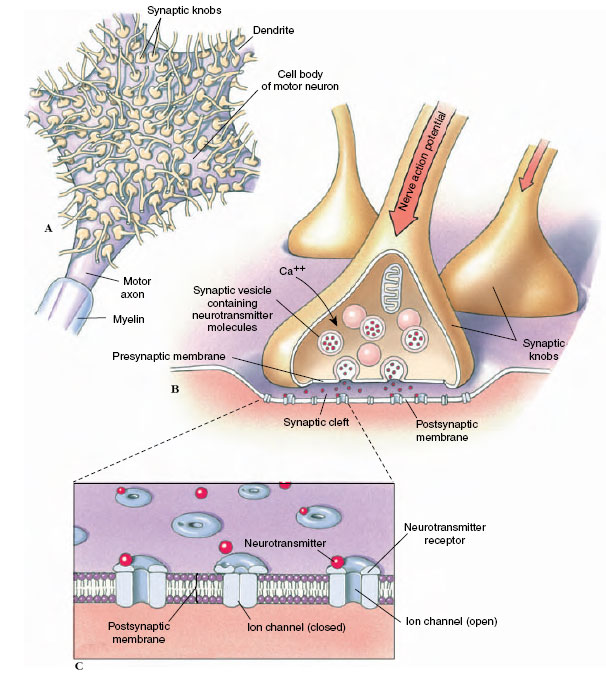 |
| Figure 35-7 Transmission of impulses across nerve synapses. A, A cell body of a motor nerve is shown with the terminations of interneurons. Each termination ends in a synaptic knob; thousands of synaptic knobs may rest on a single nerve cell body and its dendrites. B, A synaptic knob enlarged 60 times more than in A. An impulse traveling down the axon causes movement of synaptic vesicles to the presynaptic membrane where exocytosis occurs, releasing neurotransmitter molecules into the cleft. C, Diagram of a synaptic cleft at the ultrastructural level. Upon vesicular exocytosis, neurotransmitter molecules move rapidly across the gap to bind briefly with receptor molecules in the postsynaptic membrane. Binding of neurotransmitter to receptor produces a change in the potential of the postsynaptic membrane. |
The axon of most neurons divides at its end into many branches, each of which bears a synaptic knob that sits on the dendrites or cell body of the next neuron (Figure 35-7A). Because a single impulse coming down a nerve axon is transmitted along these many branches and synaptic endings on the next neuron, many impulses converge on the cell body at one instant. In addition, the axon terminations of many neurons may almost cover a nerve cell body and its dendrites with thousands of synapses. The 20 nm fluid-filled gap between presynaptic and postsynaptic membranes prevents action potentials from spreading directly to the postsynaptic neuron. Instead the synaptic knobs secrete a specific neurotransmitter that communicates chemically with the postsynaptic cell. One of the most common neurotransmitters of the peripheral nervous system is acetylcholine, which illustrates typical synaptic transmission. Inside the synaptic knobs are numerous tiny synaptic vesicles, each containing several thousand molecules of acetylcholine. Evidence suggests that when an impulse arrives at a terminal knob a sequence of events occurs as portrayed in Figure 35-8. The action potential causes an inward movement of calcium (Ca+) ions through channels in the synaptic knob membrane and this induces exocytosis of some neurotransmitter-filled synaptic vesicles. Acetylcholine molecules diffuse across the gap in a fraction of a millisecond and bind briefly to receptor molecules on ion channels in the postsynaptic membrane. This creates a voltage change in the postsynaptic membrane. Whether the voltage change is large enough to trigger a postsynaptic potential depends on how many acetylcholine molecules are released and how many channels are opened. Acetylcholine is rapidly destroyed by the enzyme acetylcholinesterase, which converts acetylcholine into acetate and choline. If not inactivated in this way, the neurotransmitter would continue to stimulate indefinitely. Organophosphate insecticides (such as malathion) and certain military nerve gases are poisonous for precisely this reason; they block acetylcholinesterase. The final step in the sequence is reabsorption of choline into the presynaptic terminal, resynthesis of acetylcholine and its storage in synaptic vesicles, ready to respond to another impulse.
Many different chemical neurotransmitters have been identified in both vertebrate and invertebrate nervous systems. Some, such as acetylcholine, norepinephrine, and glutamate, depolarize postsynaptic membranes; they are released at excitatory synapses. Other neurotransmitters, such as gamma aminobutyric acid (GABA), hyperpolarize postsynaptic membranes; thereby stabilizing them against depolarization. These neurotransmitters are released at inhibitory synapses. Neurons in the central nervous system have both excitatory and inhibitory synapses among the hundreds or thousands of synaptic knobs on the dendrites and cell body of each neuron.
The net balance of all excitatory and inhibitory inputs received by a postsynaptic cell determines whether it generates an action potential (Figure 35-8). If many excitatory impulses are received at one time, they can reduce the resting membrane potential enough in the postsynaptic membrane to elicit an action potential. Inhibitory impulses, however, stabilize the postsynaptic membrane, making it less likely that an action potential will be generated. The synapse is a crucial part of the decision-making equipment of the central nervous system, modulating flow of information from one neuron to the next.
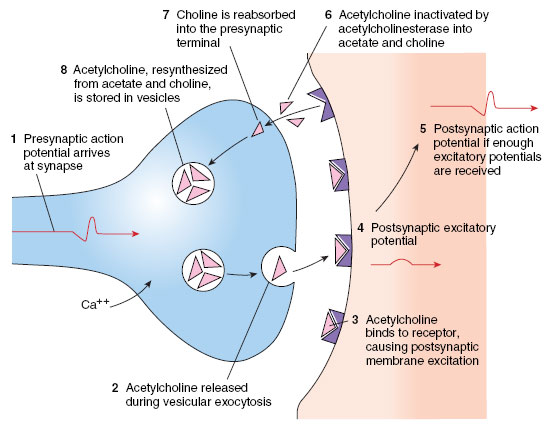 |
| Figure 35-8 Sequence of events in synaptic transmission. |
Support our developers




