Initiation of DNA replication

Fig. 26.17. Model of the initiation of DNA replication at oriC of E. coli.
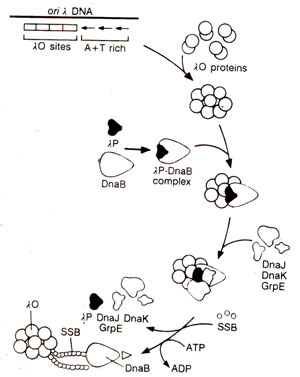
Fig. 26.18. Model of the initiation of DNA replication at oriλof phage λ.
For initiation from oriC, following steps are involved (as deduced from in vitro studies :
(i) DnaA-ATP complex (10-20 monomers of DnaA) binds at 9bp inverted repeat regions (R1, R2, R3, R4)of oriC and promotes opening of the DNA in a region of three direct repeats of a 13-bp sequence (called 13-mers; see Fig. 26.17). The opening occurs from right 13-mer leftwards, and requires negatively supercoiled DNA and Hu or IHF proteins, besides DnaA-ATP complex,
(iii) 'DnaB helicase' causes unwinding of the DNA, on addition of ATP, SSB (single strand binding protein) and DNA gyrase. This leads to the formation of a preprinting complex. Unwinding and replication from oriC proceeds in both directions (bidirectional), SSB binding occurs on single stranded regions, and two DnaB complexes, are loaded one on each strand,
(iv) Unwinding is followed by the synthesis of RNA primers by DnaG primase (through its interaction with DnaB) and the primers are elongated by 'DNA Pol III HE' (HE = holoenzyme). However, in vivo, priming of replication, in E. coli DNA, X DNA and in the leading DNA strand of Col El, is dependent on synthesis of RNA primer by RNA polymerase. (The mobile complex of helicase and primase has been termed a 'primosome'). On both, oriC and oriPl, DnaA and DnaC proteins help in the transfer of DnaB. However in λphage, its own specific proteins λO and λP are available, along with many E. coli proteins, for replication. λOand λP are analogous to E. coli's DnaA and DnaC respectively. Activation of DnaB in λphage requires release of λP, which is assisted by three heat-shock proteins, DnaK, DnaJ and GrpE (Fig. 26.18). Loading of DnaB on ColEl is dependent on PriA and Φ X type primosome.
Fig. 26.17. Model of the initiation of DNA replication at oriC of E. coli.
Fig. 26.18. Model of the initiation of DNA replication at oriλof phage λ.
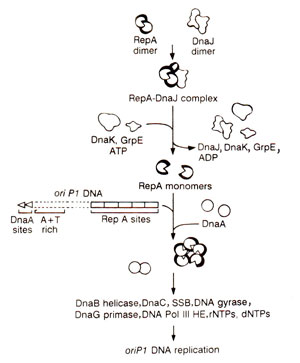
Fig. 26.19. Model of the initiation of DNA replication of oriP1 of phage P1.
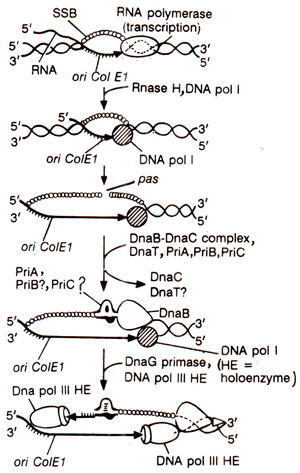
Fig. 26.20. Model of the initiation of DNA replication at ColE1/pBR322 origin.
Initiation of DNA replication from ColE1 origin is catalyzed by E. coli proteins, but differs in several essential features. On the leading strand RNA polymerase synthesizes the transcript forming RNA-DNA hybrid, that is processed by RNase H generating 3' OH group, which serves as a primer for elongation by DNA polymerase I forming DNA-DNA hybrid and displacing the other strand of the plasmid DNA.
Initiation of replication in single stranded DNA phages. In many cases (including phage M13, which is single stranded and mtDNA) it is the RNA polymerase, which is involved in priming the DNA synthesis through the synthesis of small RNA segment, which precedes DNA synthesis (as earlier discussed). However, in phage G4 and in phage Φ X 174 (both single stranded DNA), in place of RNA polymerase, a much smaller protein called primase (coded by dnaG gene) is utilized. This primase enzyme synthesizes RNA from one unique specific site each in phages G4 and M13, but at multiple sites in Φ X 174.
Fig. 26.19. Model of the initiation of DNA replication of oriP1 of phage P1.
Fig. 26.20. Model of the initiation of DNA replication at ColE1/pBR322 origin.
Support our developers




