Terpenoid Indole Alkaloid Biosynthesis
Two steps in secologanin biosynthesis are catalyzed by P450-dependent enzymes: geraniol 10-hydroxylase (G10H) converts geraniol to 10-hydroxygeraniol (Collu et al., 2001), while secologanin synthase (CYP72A1) converts loganin to secologanin and showsepidermis-specific expression in immature leaves of C. roseus (Irmler et al., 2000). The supply of terpenoid precursors should be rate-limiting in terpenoid indole alkaloid biosynthesis. The addition of secologanin or loganin to C. roseus cell culture increases alkaloid accumulation (Whitmer et al., 1998), and the
STR is a key enzyme in terpenoid indole alkaloid biosynthesis and cDNAs have been isolated from Rauvolfia serpentina (Kutchan et al., 1988) and C. roseus (Mcknight et al., 1990). STR is one of the most investigated biosynthetic genes in secondary metabolism. Strictosidine is deglucosylated by strictosidine glucosidase (SGD) (Geerlings et al., 2000) and then converted via several unstable intermediates. While there is limited information available on the pathway to catharanthine, vindoline biosynthesis has been relatively well characterized, although the production of vindoline in cultured cells is limited.
The first of six steps in the conversion of tabersonine to vindoline consists of hydroxylation at the C-16 position by tabersonine 16-hydroxylase (T16H), a P450-dependent monooxygenase. While several P450 sequences
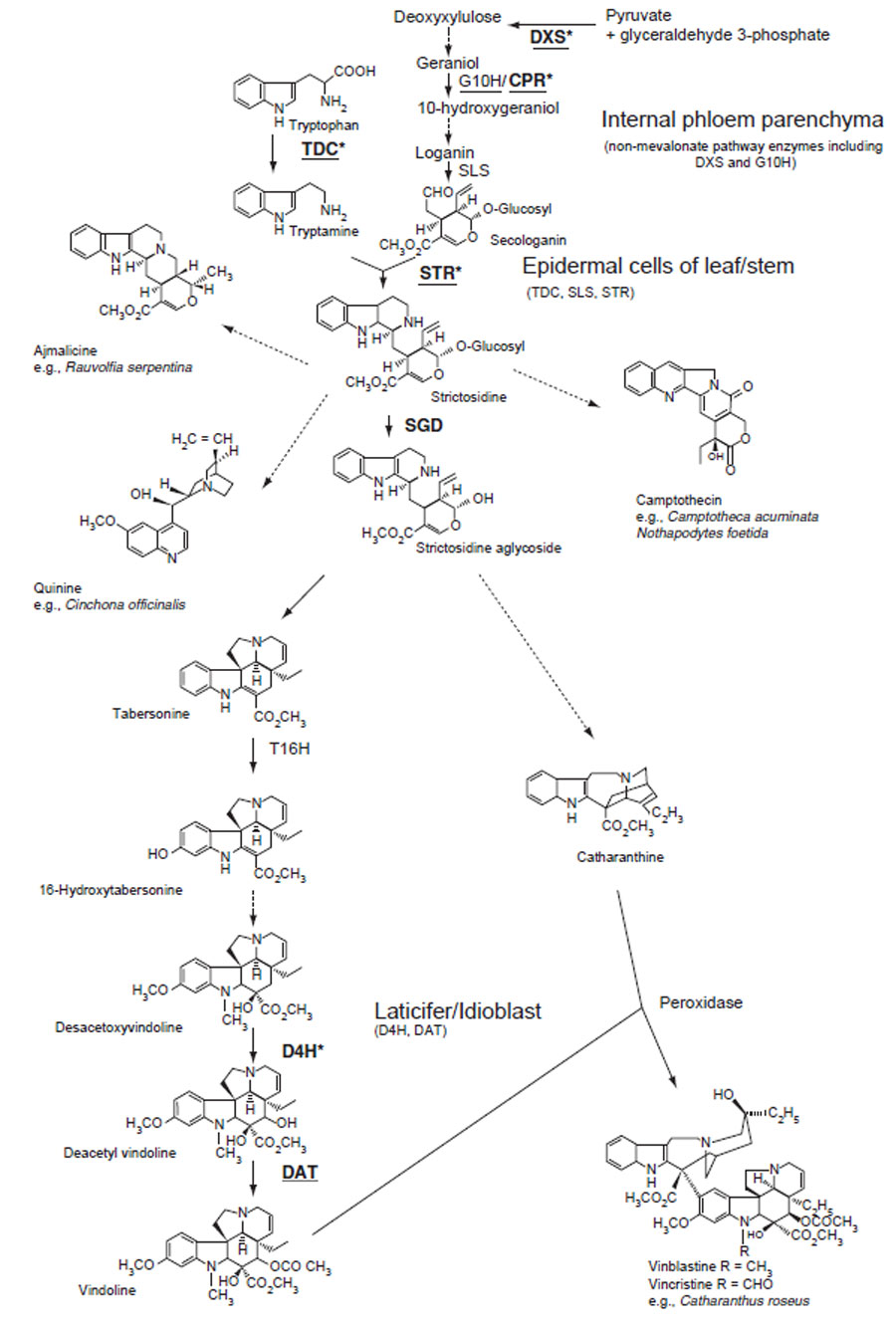 |
| FIGURE 11.3 Biosynthetic pathways to various terpenoid indole alkaloids. Unbroken arrows indicate single enzymatic conversions and dotted arrows indicate multiple enzymatic steps. Enzymes for which the corresponding genes have been cloned are indicated. JA-inducible genes are indicated in bold. Underlining indicates that the corresponding genes have been tested for being an ORCA3 target gene in C. roseus cells. Enzyme-encoding genes regulated by ORCA3 are asterisked (Vazquez-Flota et al., 2000). DXS, D-1-deoxyxylulose 5-phosphate synthase; STR, strictosidine synthase; TDC, tryptophan decarboxylase; G10H, geraniol 10-hydroxylase; CYP72A1, secologanin synthase; SGD, strictosidine glucosidase; T16H, tabersonine 16-hydroxylase; CPR, cytochrome P450 reductase; D4H, desacetoxyvindoline 4-hydroxylase; DAT, acetylcoenzyme A: deacetylvindoline 4-O-acetyltransferase. |
The penultimate step in vindoline biosynthesis is catalyzed by a cytosolic 2-oxoglutarate-dependent dioxygenase that hydroxylates the C-4 position of desacetoxyvindoline 4-hydroxylase (D4H) (Vazquez-Flota et al., 1997), and the final step is catalyzed by the cytosolic acetylcoenzyme A: deacetylvindoline 4-Oacetyltransferase (DAT) (St-Pierre et al., 1998). The expression of T16H, D4H, and DAT in developing C. roseus seedlings is light regulated. Although D4H and DAT activities are detected exclusively under conditions that result in vindoline biosynthesis, T16H is expressed at low levels in C. roseus cell cultures that do not accumulate vindoline (St-Pierre and De Luca, 1995). The expression of D4H appears to be under complex, multilevel developmental and light regulation.
A series of experiments with leaves of C. roseus (Burlat et al., 2004; St-Pierre et al., 1999) showed that at least three cell types are involved in vindoline biosynthesis.
Vindoline biosynthesis is restricted to the aboveground organs, and the pathway beyond tabersonine is not expressed in tissue cultures (Vazquez-Flota et al., 2002), whereas catharanthine accumulates in cultured cells as well as etiolated seedlings.
Until recently, characterization of terpenoid indole alkaloid biosynthesis has mainly been carried out with C. roseus, but the recent establishment of hairy root cultures of Ophiorrhiza pumila (Rubiaceae) that showed high camptothecin production provided another useful experimental system. Computer-aided atomic reconstruction of metabolism and tracer experiments with [1–13C] glucose indicated that camptothecin is formed by the combined activities of the 2C-methyl-D-erythritol 4-phosphate pathway and the shikimate pathway (Yamazaki et al., 2004).
Support our developers




