Tropane Alkaloid and Nicotine Biosynthesis
Tropinone is located at a branch point in tropane alkaloid synthesis. Two related dehydrogenases, tropinone reductase I (TR-I) and tropinone reductase II (TR-II), stereospecifically reduce the 3-keto group of tropinone to the 3α- and 3β-groups of tropine and ψ-tropine, respectively. cDNA clones for TR-I and TR-II have been isolated from Datura stramonium (Nakajima et al., 1993). A further analysis of their localization suggested that TR-I and TR-II were localized differently and might have different functions (Nakajima and Hashimoto, 1999). Nortropane polyhydroxylated alkaloids, calistegines, are also assumed to originate from ψ-tropine. They have been isolated from different species in the Solanaceae (Scholl et al.,
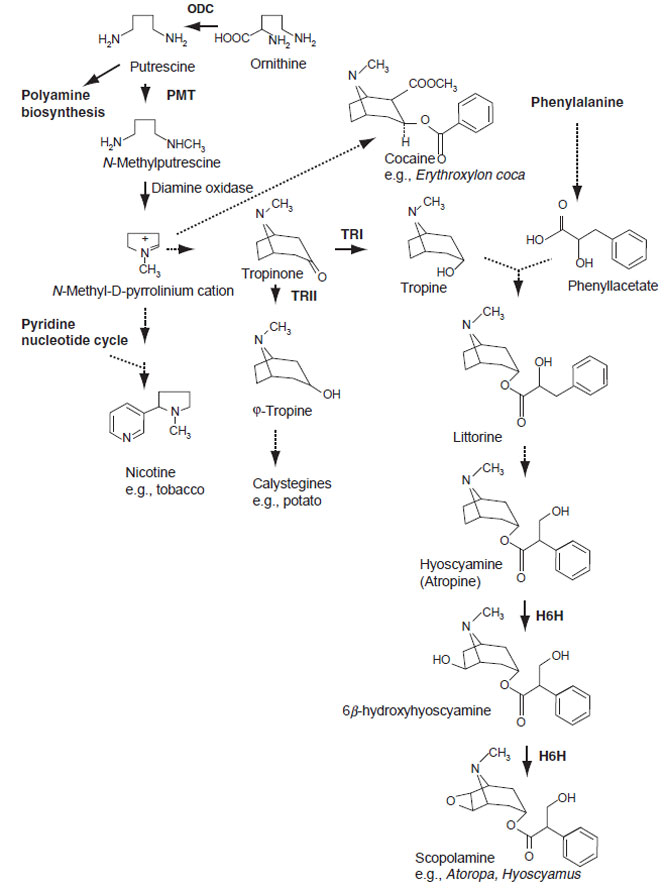 |
| FIGURE 11.4 Biosynthetic pathways to tropane alkaloids, related compounds, and nicotine. Unbroken arrows indicate single enzymatic conversions and broken arrows indicate multiple enzymatic steps. Enzymes for which the corresponding genes have been cloned are indicated in bold. ODC, ornithine decarboxylase; PMT, putrescine N-methyltransferase; TR-I/II, tropinone reductase I/II; H6H, hyoscyamine 6β-hydroxylase. |
Hyoscyamine is produced by condensation of tropine and the phenylalaninederived intermediate (R)-phenyllactate. Hyoscyamine can be converted to its epoxide scopolamine via 6β-hydroxylhyoscyamine by a 2-oxoglutarate-dependent dioxygenase, hyoscyamine 6β-hydroxylase (H6H) (Matsuda et al., 1991). H6H localizes in the pericycle in branch roots of several scopolamine-producing Solanaceae plants (Hashimoto et al., 1991). Histochemical analysis using H. niger and A. belladonna H6H promoter::GUS fusion gene also showed that cell-specific expression of the H6H gene is controlled by (unknown) genetic regulation specific to scopolamine-producing plants but is absent in tobacco that does not produce scopolamine (Kanegae et al., 1994; Suzuki et al., 1999b).
In Nicotiana sylvestris, a set of nicotine biosynthesis genes was activated by the exogenous application of methyl jasmonate (MeJA), but this activation was effectively suppressed by simultaneous treatment with ethylene (Shoji et al., 2000), even though ethylene and JA are generally considered to act synergistically. In contrast, treatment of A. belladonna roots with MeJA did not lead to upregulated expression of AbPMT genes (Suzuki et al., 1999a). The different responses of tropane alkaloids and nicotine biosynthesis to JA and ethylene suggest that these biosyntheses might be under the control of different genetic regulation systems (Shoji et al., 2000).
Support our developers




