Components of Eukaryotic Cells and Their Functions
Components of Eukaryotic
Cells and Their Functions
Typically, eukaryotic cells are enclosed
within a thin, selectively permeable cell membrane (Figure 3-4). The most
prominent organelle is the spherical or
ovoid nucleus, enclosed within two
membranes to form the double-layered nuclear envelope (Figure 3-4). The
region outside the nucleus is regarded
as cytoplasm. Within the cytoplasm
are many organelles, such as mitochondria,
Golgi complexes, centrioles,
and endoplasmic reticulum.
Plant cells typically contain plastids,
some of which are photosynthetic
organelles, and plant cells bear a cell
wall containing cellulose outside the
cell membrane.
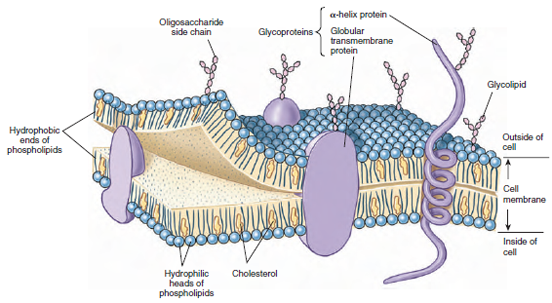
The fluid-mosaic model is the currently accepted concept of cell membranes. By electron microscopy, the cell membrane appears as two dark lines, each approximately 3 nm thick, at each side of a light zone (Figure 3-5). The entire membrane is 8 to 10 nm thick. This image is the result of a phospholipid bilayer, two layers of phospholipid molecules, all oriented with their water-soluble ends toward the outside and their fat-soluble portions toward the inside of the membrane (Figure 3-6). An important characteristic of the phospholipid bilayer is that it is liquid, giving the membrane flexibility and allowing the phospholipid molecules to move sideways freely within their own monolayer. Molecules of cholesterol are interspersed in the lipid portion of the bilayer (Figure 3-6). They make the membrane even less permeable and decrease its flexibility.
Glycoproteins (proteins with carbohydrates attached) are essential components of cell membranes. Some of these proteins catalyze the transport of substances such as negatively charged ions across the membrane. Others act as specific receptors for various molecules or as highly specific markings. For example, the self/nonself recognition that enables the immune system to react to invaders (Immunity) is based on proteins of this type. Some aggregations of protein molecules form pores through which small polar molecules may enter. Like the phospholipid molecules, most of the glycoproteins can move laterally in the membrane, although more slowly.
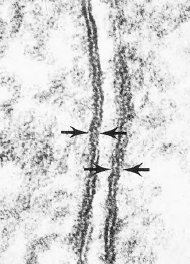
Nuclear envelopes contain less
cholesterol than cell membranes, and
pores in the envelope (Figure 3-7)
allow molecules to move between
nucleus and cytoplasm. Nuclei contain
chromatin, a complex of DNA,
basic proteins called histones, and
nonhistone protein. Chromatin carries
the genetic information, the code that
results in most of the components
characteristic of the cell after transcription
and translation (see Principles of Genetics:A Review). Nucleoli are specialized parts of
certain chromosomes that stain in a
characteristically dark manner. They
carry multiple copies of the DNA
information to synthesize ribosomal
RNA. After transcription from DNA,
ribosomal RNA combines with protein
to form a ribosome, detaches from
the nucleolus, and passes to the cytoplasm
through pores in the nuclear
envelope.
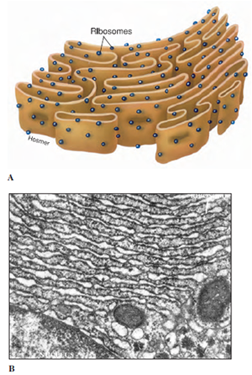
The outer membrane of the nuclear envelope is continuous with extensive membranous elements in the cytoplasm called endoplasmic reticulum (ER) (Figures 3-7 and 3-8). The space between the membranes of the nuclear envelope communicates with channels (cisternae) in the ER. The ER is a complex of membranes that separates some of the products of the cell from the synthetic machinery that produces them, apparently functioning as routes for transport of proteins within the cell. Membranes of the ER may be covered on their outer surfaces with ribosomes and are thus designated rough ER, or they may lack ribosomal covering and be called smooth ER. Smooth ER functions in synthesis of lipids and phospholipids. Protein synthesized by ribosomes on rough ER enters the cisternae and from there is transported to the Golgi apparatus or complex.
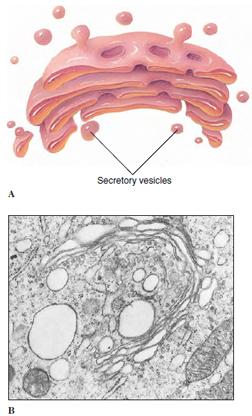
The Golgi complex (Figures 3-9
and 3-10) is composed of a stack of
membranous vesicles that function in
storage, modification, and packaging of
protein products, especially secretory
products. The vesicles do not synthesize
protein but may add complex carbohydrates
to the molecules. Small vesicles
of ER containing protein pinch off
and then fuse with sacs on the “forming
face” of a Golgi complex. After modification,
the proteins bud off vesicles on
the “maturing face” of the complex (Figure
3-10). The contents of some of
these vesicles may be expelled to the
outside of the cell, as secretory products
destined to be exported from a glandular
cell. Others may contain digestive
enzymes that remain in the same cell
that produces them. Such vesicles are
called lysosomes (literally “loosening
body,” a body capable of causing lysis,
or disintegration). Enzymes that they
contain are involved in the breakdown
of foreign material, including bacteria
engulfed by the cell. Lysosomes also are
capable of breaking down injured or
diseased cells and worn-out cellular
components. Their enzymes are so
powerful that they kill the cell that
formed them if the lysosome membrane
ruptures. In normal cells the enzymes
remain safely enclosed within the protective
membrane. Lysosomal vesicles
may pour their enzymes into a larger
membrane-bound body containing an
ingested food particle, the food vacuole
or phagosome. Other vacuoles,
such as contractile vacuoles of some
single-celled organisms, may
contain only fluid and function to regulate
ions and water.
Mitochondria (sing., mitochondrion)
(Figure 3-11) are conspicuous
organelles present in nearly all eukaryotic
cells. They are diverse in size, number,
and shape; some are rodlike, and
others are more or less spherical. They
may be scattered uniformly through the
cytoplasm, or they may be localized
near cell surfaces and other regions
where there is high metabolic activity. A
mitochondrion is composed of a double
membrane. The outer membrane is
smooth, whereas the inner membrane is
folded into numerous platelike or fingerlike
projections called cristae (Figure
3-11), which increases internal surface
area where chemical reactions take
place. These characteristic features
make mitochondria easy to identify
among the organelles. Mitochondria are
often called “powerhouses of the cell,”
because enzymes located on the cristae
carry out the energy-yielding steps of
aerobic metabolism. ATP (adenosine triphosphate),
the most important energytransfer
molecule of all cells, is produced
in this organelle. Mitochondria
are self-replicating. They have a tiny,
circular genome, much like the
genomes of prokaryotes except that it is
much smaller. It contains DNA that
specifies some, but not all, of the proteins
of the mitochondrion.
Eukaryotic cells characteristically have a system of tubules and filaments that form the cytoskeleton (Figures 3-12 and 3-13). These provide support and maintain the form of cells, and in many cells, they provide a means of locomotion and translocation of organelles within the cell. Microfilaments are thin, linear structures, first observed distinctly in muscle cells, where they are responsible for the ability of the cell to contract. They are made of a protein called actin. Several dozen other proteins are known that bind with actin and determine its configuration and behavior in particular cells. One of these is myosin, whose interaction with actin causes contraction in muscle and other cells. Actin microfilaments also provide a means for moving messenger RNA from the nucleus to particular positions within the cell. Microtubules, somewhat larger than microfilaments, are tubular structures composed of a protein called tubulin (Figure 3-13). They play a vital role in moving the chromosomes toward the daughter cells during cell division as will be seen later, and they are important in intracellular architecture, organization, and transport. In addition, microtubules form essential parts of the structures of cilia and flagella. Microtubules radiate out from a microtubule organizing center, the centrosome, near the nucleus.
Centrosomes
are not membrane bound. Within centrosomes
are found a pair of centrioles
(Figures 3-4 and 3-14), which are
themselves composed of microtubules.
Microtubules radiating from
the centrioles form the aster. Each
centriole of a pair lies at right angles
to the other and is a short cylinder of
nine triplets of microtubules. They
replicate before cell division. Although
cells of higher plants do not
have centrioles, a microtubule organizing
center is present. Intermediate
filaments are larger than microfilaments
but smaller than microtubules.
There are five biochemically distinct
types of intermediate filaments, and
their composition and arrangement
depend on the cell type in which they
are found.
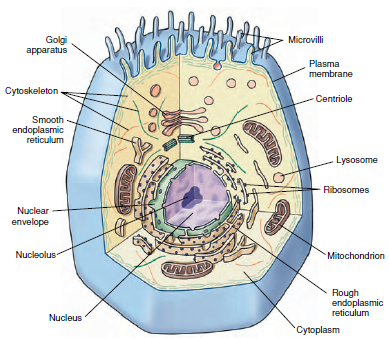 |
| Figure 3-4 Generalized cell with principal organelles, as might be seen with the electron microscope. No single cell contains all these organelles, but many cells contain a large number of them. |
The fluid-mosaic model is the currently accepted concept of cell membranes. By electron microscopy, the cell membrane appears as two dark lines, each approximately 3 nm thick, at each side of a light zone (Figure 3-5). The entire membrane is 8 to 10 nm thick. This image is the result of a phospholipid bilayer, two layers of phospholipid molecules, all oriented with their water-soluble ends toward the outside and their fat-soluble portions toward the inside of the membrane (Figure 3-6). An important characteristic of the phospholipid bilayer is that it is liquid, giving the membrane flexibility and allowing the phospholipid molecules to move sideways freely within their own monolayer. Molecules of cholesterol are interspersed in the lipid portion of the bilayer (Figure 3-6). They make the membrane even less permeable and decrease its flexibility.
|
|
Glycoproteins (proteins with carbohydrates attached) are essential components of cell membranes. Some of these proteins catalyze the transport of substances such as negatively charged ions across the membrane. Others act as specific receptors for various molecules or as highly specific markings. For example, the self/nonself recognition that enables the immune system to react to invaders (Immunity) is based on proteins of this type. Some aggregations of protein molecules form pores through which small polar molecules may enter. Like the phospholipid molecules, most of the glycoproteins can move laterally in the membrane, although more slowly.
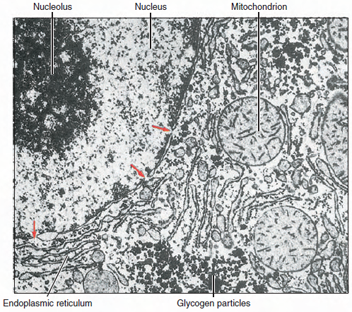 |
| Figure 3-7 Electron of part of hepatic cell of rat showing portion of nucleus (left) and surrounding cytoplasm. Endoplasmic reticulum and mitochondria are visible in cytoplasm, and pores (arrows) can be seen in nuclear envelope. (× 14,000) |
The outer membrane of the nuclear envelope is continuous with extensive membranous elements in the cytoplasm called endoplasmic reticulum (ER) (Figures 3-7 and 3-8). The space between the membranes of the nuclear envelope communicates with channels (cisternae) in the ER. The ER is a complex of membranes that separates some of the products of the cell from the synthetic machinery that produces them, apparently functioning as routes for transport of proteins within the cell. Membranes of the ER may be covered on their outer surfaces with ribosomes and are thus designated rough ER, or they may lack ribosomal covering and be called smooth ER. Smooth ER functions in synthesis of lipids and phospholipids. Protein synthesized by ribosomes on rough ER enters the cisternae and from there is transported to the Golgi apparatus or complex.
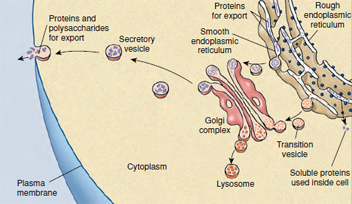 |
| Figure 3-10 System for assembling, isolating, and secreting proteins for export in a eukaryotic cell. |
|
|
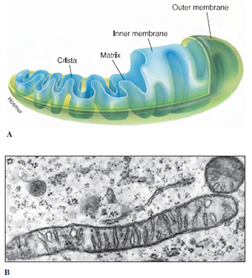 |
| Figure 3-11 Mitochondria. A, Structure of a typical mitochondrion. B, Electron micrograph of mitochondria in cross and longitudinal section. (×30,000) |
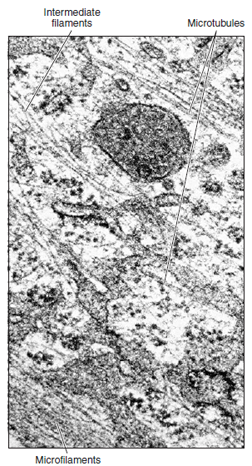 |
| Figure 3-12 Cytoskeleton of a cell, showing its complex nature. Three visible cytoskeletal elements, in order of increasing diameter, are microfilaments, intermediate filaments, and microtubules (×66,600) |
Eukaryotic cells characteristically have a system of tubules and filaments that form the cytoskeleton (Figures 3-12 and 3-13). These provide support and maintain the form of cells, and in many cells, they provide a means of locomotion and translocation of organelles within the cell. Microfilaments are thin, linear structures, first observed distinctly in muscle cells, where they are responsible for the ability of the cell to contract. They are made of a protein called actin. Several dozen other proteins are known that bind with actin and determine its configuration and behavior in particular cells. One of these is myosin, whose interaction with actin causes contraction in muscle and other cells. Actin microfilaments also provide a means for moving messenger RNA from the nucleus to particular positions within the cell. Microtubules, somewhat larger than microfilaments, are tubular structures composed of a protein called tubulin (Figure 3-13). They play a vital role in moving the chromosomes toward the daughter cells during cell division as will be seen later, and they are important in intracellular architecture, organization, and transport. In addition, microtubules form essential parts of the structures of cilia and flagella. Microtubules radiate out from a microtubule organizing center, the centrosome, near the nucleus.
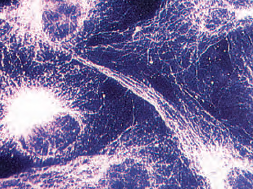 |
| Figure 3-13 The microtubules in kidney cells of a baby hamster have been rendered visible by treatment with a preparation of fluorescent proteins that specifically bind to tubulin. |
Support our developers




