Phases in Mitosis
Phases in Mitosis
There are two distinct stages of cell division: division of the nuclear chromosomes (mitosis) and division of the cytoplasm (cytokinesis). Mitosis (that is, chromosomal segregation) is certainly the most obvious and complex part of cell division and that of greatest interest to the cytologist. Cytokinesis normally immediately follows mitosis, although occasionally the nucleus may divide a number of times without a corresponding division of the cytoplasm. In such a case the resulting mass of protoplasm containing many nuclei is referred to as a multinucleate cell. An example is the giant resorptive cell type of bone (osteoclast), which may contain 15 to 20 nuclei. Sometimes a multinucleate mass is formed by cell fusion rather than nuclear proliferation. This arrangement is called a syncytium. An example is vertebrate skeletal muscle, which is composed of multinucleate fibers formed by the fusion of numerous embryonic cells.
The process of mitosis is divided into four successive stages or phases, although one stage merges into the next without sharp lines of transition. These phases are prophase, metaphase, anaphase, and telophase (Figure 3-22). When cells are not actively dividing, they are in interphase, during which DNA replicates and genes are transcribed.
Prophase
At the beginning of prophase, the centrosomes (along with their centrioles) replicate, and the two centrosomes migrate to opposite sides of the nucleus (Figure 3-22). At the same time, microtubules appear between the two centrosomes to form a football-shaped spindle, so named because of its resemblance to nineteenth-century wooden spindles, used to twist thread together in spinning. Other microtubules radiate outward from each centrosome to form asters.
At this time the diffuse nuclear chromatin condenses to form visible chromosomes. These actually consist of two identical sister chromatids formed during interphase. The sister chromatids are joined together at their centromere. Dynamic spindle fibers repeatedly extend and retract from the centrosome. When a fiber encounters a kinetochore, it binds to the kinetochore, ceases extending and retracting, and is now called a kinetochore fiber. It is as if centrosomes send out “feelers” to find chromosomes.
Microtubules are long, hollow, inelastic
cylinders composed of the protein tubulin
(Figure 3-23). Each tubulin molecule is actually
a doublet composed of two globular
proteins.The molecules are attached headto-
tail to form a strand, and 13 strands
aggregate to form a microtubule. Because
the tubulin subunits in a microtubule are
always attached head-to-tail, the ends of the
microtubule differ chemically and functionally.
One end (called the plus end) both
adds and deletes tubulin subunits more
rapidly than the other end (the minus end).
In a mitotic spindle, the plus ends of the
kinetochore and polar fibers are away from
the centrosome, and the minus ends are at
the centrosome.The microtubule grows
when the rate of adding subunits exceeds
that of removing them, and it becomes
shorter when the rate of removal exceeds
that of addition.
Metaphase
Each centromere has two kinetochores, and each of the kinetochores is attached to one of the centrosomes by a kinetochore fiber. By a kind of tugof- war during metaphase, the condensed sister chromatids are moved to the middle of the nuclear region to form a metaphasic plate (Figure 3- 24). The centromeres line up precisely on the plate with the arms of the chromatids trailing off randomly in various directions.
Anaphase
The single centromere that has held the two chromatids together now splits so that two independent chromosomes, each with its own centromere, are formed. The chromosomes move toward their respective poles, pulled by the kinetochore fibers. This phase is often called anaphase A. The arms of each chromosome trail along behind as the microtubules shorten to drag the chromosomes along. Present evidence indicates that the force moving the chromosomes is disassembly of the tubulin subunits at the kinetochore end of the microtubules.
As the chromosomes approach their respective centrosomes, the spindle lengthens, and the centrosomes move farther apart. This is anaphase B. The mechanism of this movement appears to involve the interdigitating free ends of the polar fibers. Tubulin in these microtubules has other protein molecules associated with it that serve as “motor molecules.” These motor molecules interact with the adjacent fiber (or motor molecules on the adjacent fiber) and push the two halves of the spindle away from each other.
Telophase
When daughter chromosomes reach their respective poles, telophase has begun. Daughter chromosomes are crowded together and stain intensely with histological stains. Spindle fibers disappear and chromosomes lose their identity, reverting to a diffuse chromatin network characteristic of an interphase nucleus. Finally, nuclear membranes reappear around the two daughter nuclei.
There are two distinct stages of cell division: division of the nuclear chromosomes (mitosis) and division of the cytoplasm (cytokinesis). Mitosis (that is, chromosomal segregation) is certainly the most obvious and complex part of cell division and that of greatest interest to the cytologist. Cytokinesis normally immediately follows mitosis, although occasionally the nucleus may divide a number of times without a corresponding division of the cytoplasm. In such a case the resulting mass of protoplasm containing many nuclei is referred to as a multinucleate cell. An example is the giant resorptive cell type of bone (osteoclast), which may contain 15 to 20 nuclei. Sometimes a multinucleate mass is formed by cell fusion rather than nuclear proliferation. This arrangement is called a syncytium. An example is vertebrate skeletal muscle, which is composed of multinucleate fibers formed by the fusion of numerous embryonic cells.
The process of mitosis is divided into four successive stages or phases, although one stage merges into the next without sharp lines of transition. These phases are prophase, metaphase, anaphase, and telophase (Figure 3-22). When cells are not actively dividing, they are in interphase, during which DNA replicates and genes are transcribed.
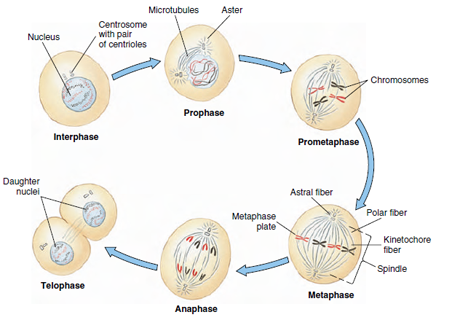 |
| Figure 3-22 Stages of mitosis, showing division of a cell with two pairs of chromosomes. One chromosome of each pair is shown in red. |
Prophase
At the beginning of prophase, the centrosomes (along with their centrioles) replicate, and the two centrosomes migrate to opposite sides of the nucleus (Figure 3-22). At the same time, microtubules appear between the two centrosomes to form a football-shaped spindle, so named because of its resemblance to nineteenth-century wooden spindles, used to twist thread together in spinning. Other microtubules radiate outward from each centrosome to form asters.
At this time the diffuse nuclear chromatin condenses to form visible chromosomes. These actually consist of two identical sister chromatids formed during interphase. The sister chromatids are joined together at their centromere. Dynamic spindle fibers repeatedly extend and retract from the centrosome. When a fiber encounters a kinetochore, it binds to the kinetochore, ceases extending and retracting, and is now called a kinetochore fiber. It is as if centrosomes send out “feelers” to find chromosomes.
 |
| Figure 3-23 A microtubule is composed of 13 strands of tubulin molecules, and each molecule is a dimer. Tubulin dimers are added to and removed from the (+) end of the microtubule more rapidly than at the (−) end. |
Metaphase
Each centromere has two kinetochores, and each of the kinetochores is attached to one of the centrosomes by a kinetochore fiber. By a kind of tugof- war during metaphase, the condensed sister chromatids are moved to the middle of the nuclear region to form a metaphasic plate (Figure 3- 24). The centromeres line up precisely on the plate with the arms of the chromatids trailing off randomly in various directions.
Anaphase
The single centromere that has held the two chromatids together now splits so that two independent chromosomes, each with its own centromere, are formed. The chromosomes move toward their respective poles, pulled by the kinetochore fibers. This phase is often called anaphase A. The arms of each chromosome trail along behind as the microtubules shorten to drag the chromosomes along. Present evidence indicates that the force moving the chromosomes is disassembly of the tubulin subunits at the kinetochore end of the microtubules.
As the chromosomes approach their respective centrosomes, the spindle lengthens, and the centrosomes move farther apart. This is anaphase B. The mechanism of this movement appears to involve the interdigitating free ends of the polar fibers. Tubulin in these microtubules has other protein molecules associated with it that serve as “motor molecules.” These motor molecules interact with the adjacent fiber (or motor molecules on the adjacent fiber) and push the two halves of the spindle away from each other.
Telophase
When daughter chromosomes reach their respective poles, telophase has begun. Daughter chromosomes are crowded together and stain intensely with histological stains. Spindle fibers disappear and chromosomes lose their identity, reverting to a diffuse chromatin network characteristic of an interphase nucleus. Finally, nuclear membranes reappear around the two daughter nuclei.
Support our developers




