Surfaces of Cells and Their Specializations
Surfaces of Cells and Their
Specializations
The free surface of epithelial cells
(cells that cover the surface of a structure
or line a tube or cavity) sometimes
bears either cilia or flagella
(sing., cilium, flagellum). These are
motile extensions of the cell surface
that sweep materials past the cell. In
many single-celled organisms and
some small multicellular forms, they
propel the entire organism through a
liquid medium. Flagella provide the
means of locomotion for male reproductive
cells of most animals and
many plants.
Cilia and flagella have different beating patterns, but their internal structure is the same. With few exceptions, the internal structures of locomotory cilia and flagella are composed of a long cylinder of nine pairs of microtubules enclosing a central pair (see Figure 11-3). At the base of each cilium or flagellum is a basal body (kinetosome), which is identical in structure to a centriole.
Indeed, cilia and flagella are so alike in details of their structure that it seems highly likely that they had a common evolutionary origin.Whether their origin was the symbiosis of a spirochete-like bacterium and host cell, as suggested by Margulis, is more conjectural. Margulis and others prefer the term undulipodia to include both cilia and flagella, and it is less awkward to use one word for structures that are alike in structure and origin.However, the terms “cilia” and “flagella” are so common and widely used that the student should be familiar with them.
Many cells move neither by cilia nor flagella but by ameboid movement using pseudopodia. Some groups of protozoa , migrating cells in embryos of multicellular animals, and some cells of adult multicellular animals, such as white blood cells, show ameboid movement. Cytoplasmic streaming through the action of actin microfilaments extends a lobe (pseudopodium) outward from the surface of the cell. Continued streaming in the direction of the pseudopodium brings cytoplasmic organelles into the lobe and accomplishes movement of the entire cell. Some specialized pseudopodia have cores of microtubules, and movement is effected by assembly and disassembly of the tubular rods.
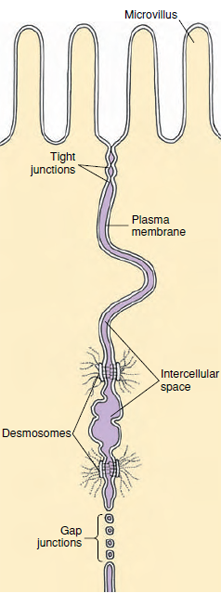
Cells covering the surface of a structure (epithelial cells) or cells packed together in a tissue may have specialized junctional complexes between them. Nearest the free surface, the membranes of two cells next to each other appear to fuse, forming a tight junction (Figure 3-15). Tight junctions function as seals to prevent the passage of molecules between cells from one side of a layer of cells to another, because there is usually a space of about 20 nm between the cell membranes of adjacent cells. Tight junctions between intestinal cells, for example, force molecules absorbed from the intestinal contents to pass through the epithelial cells, rather than between them. At various points beneath tight junctions, small ellipsoid discs occur, just within the cell membrane in each cell. These appear to act as “spot-welds” and are called desmosomes. From each desmosome a tuft of intermediate filaments extends into the cytoplasm, and linker proteins extend through the cell membrane into the intercellular space to bind the discs together. Desmosomes are not seals but seem to increase the strength of the tissue. Gap junctions, rather than serving as points of attachment, provide a means of intercellular communication. They form tiny canals between cells, so that their cytoplasm becomes continuous, and small molecules can pass from one cell to the other. Gap junctions may occur between cells of epithelial, nervous, and muscle tissues.
Another specialization of the cell surfaces is the lacing together of adjacent cell surfaces where the cell membranes of the cells infold and interdigitate very much like a zipper. They are especially common in the epithelial cells of kidney tubules. The distal or apical boundaries of some epithelial cells, as seen by electron microscopy, show regularly arranged microvilli. They are small, fingerlike projections consisting of tubelike evaginations of the cell membrane with a core of cytoplasm (Figure 3-16). They are seen clearly in the lining of the intestine where they greatly increase the absorptive and digestive surface. Such specializations appear as brush borders by light microscopy.
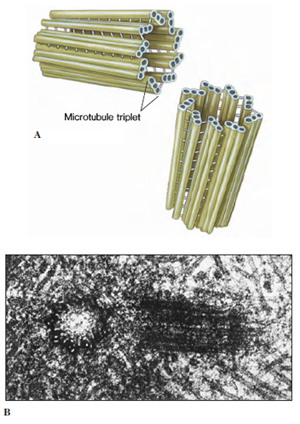 |
| Figure 3-14 Centrioles. A, Each centriole is composed of nine triplets of microtubules arranged as a cylinder. B, Electron micrograph of a pair of centrioles, one in longitudinal (right) and one in cross section (left). The normal orientation of centrioles is at right angles to each other. |
Cilia and flagella have different beating patterns, but their internal structure is the same. With few exceptions, the internal structures of locomotory cilia and flagella are composed of a long cylinder of nine pairs of microtubules enclosing a central pair (see Figure 11-3). At the base of each cilium or flagellum is a basal body (kinetosome), which is identical in structure to a centriole.
Indeed, cilia and flagella are so alike in details of their structure that it seems highly likely that they had a common evolutionary origin.Whether their origin was the symbiosis of a spirochete-like bacterium and host cell, as suggested by Margulis, is more conjectural. Margulis and others prefer the term undulipodia to include both cilia and flagella, and it is less awkward to use one word for structures that are alike in structure and origin.However, the terms “cilia” and “flagella” are so common and widely used that the student should be familiar with them.
Many cells move neither by cilia nor flagella but by ameboid movement using pseudopodia. Some groups of protozoa , migrating cells in embryos of multicellular animals, and some cells of adult multicellular animals, such as white blood cells, show ameboid movement. Cytoplasmic streaming through the action of actin microfilaments extends a lobe (pseudopodium) outward from the surface of the cell. Continued streaming in the direction of the pseudopodium brings cytoplasmic organelles into the lobe and accomplishes movement of the entire cell. Some specialized pseudopodia have cores of microtubules, and movement is effected by assembly and disassembly of the tubular rods.
Cells covering the surface of a structure (epithelial cells) or cells packed together in a tissue may have specialized junctional complexes between them. Nearest the free surface, the membranes of two cells next to each other appear to fuse, forming a tight junction (Figure 3-15). Tight junctions function as seals to prevent the passage of molecules between cells from one side of a layer of cells to another, because there is usually a space of about 20 nm between the cell membranes of adjacent cells. Tight junctions between intestinal cells, for example, force molecules absorbed from the intestinal contents to pass through the epithelial cells, rather than between them. At various points beneath tight junctions, small ellipsoid discs occur, just within the cell membrane in each cell. These appear to act as “spot-welds” and are called desmosomes. From each desmosome a tuft of intermediate filaments extends into the cytoplasm, and linker proteins extend through the cell membrane into the intercellular space to bind the discs together. Desmosomes are not seals but seem to increase the strength of the tissue. Gap junctions, rather than serving as points of attachment, provide a means of intercellular communication. They form tiny canals between cells, so that their cytoplasm becomes continuous, and small molecules can pass from one cell to the other. Gap junctions may occur between cells of epithelial, nervous, and muscle tissues.
Another specialization of the cell surfaces is the lacing together of adjacent cell surfaces where the cell membranes of the cells infold and interdigitate very much like a zipper. They are especially common in the epithelial cells of kidney tubules. The distal or apical boundaries of some epithelial cells, as seen by electron microscopy, show regularly arranged microvilli. They are small, fingerlike projections consisting of tubelike evaginations of the cell membrane with a core of cytoplasm (Figure 3-16). They are seen clearly in the lining of the intestine where they greatly increase the absorptive and digestive surface. Such specializations appear as brush borders by light microscopy.
|
|

Support our developers




