Diffusion and Osmosis
Diffusion and Osmosis
Diffusion is a movement of particles
from an area of high concentration to
an area of lower concentration of the
particles or molecules, thus tending to
equalize the concentration throughout
the area of diffusion. If a living cell surrounded
by a membrane is immersed
in a solution having a higher concentration
of solute molecules than the fluid
inside the cell, a concentration gradient instantly exists between the two
fluids. Assuming that the membrane is permeable to the solute, there is a net
movement of solute toward the inside,
the side having the lower concentration.
The solute diffuses “downhill”
across the membrane until its concentrations
on each side are equal.
Most cell membranes are selectively permeable, that is, permeable to water but variably permeable or impermeable to solutes. In free diffusion it is this selectiveness that regulates molecular traffic. As a rule, gases (such as oxygen and carbon dioxide), urea, and lipid-soluble solutes (such as hydrocarbons and alcohol) are the only solutes that can diffuse through biological membranes with any degree of freedom. Because many water-soluble molecules readily pass through membranes, such movements cannot be explained by simple diffusion. Sugars, as well as many electrolytes and macromolecules, are moved across membranes by carriermediated processes, which are described in the next section.
If we place a membrane between two unequal concentrations of solutes to which the membrane is impermeable, water flows through the membrane from the more dilute to the more concentrated solution. The water molecules move across the membrane down a concentration gradient from an area where the water molecules are more concentrated to an area on the other side of the membrane where they are less concentrated. This is osmosis.
We can demonstrate osmosis by a simple experiment in which we tie a selectively permeable membrane such as cellophane tightly over the end of a funnel. We fill the funnel with a salt solution and place it in a beaker of pure water so that the water levels inside and outside the funnel are equal. In a short time the water level in the glass tube of the funnel rises, indicating a net movement of water through the cellophane membrane into the salt solution (Figure 3-17).
Inside the funnel are salt molecules, as well as water molecules. In the beaker outside the funnel are only water molecules. Thus the concentration of water is less on the inside because some of the available space is occupied by the larger, nondiffusible salt molecules. A concentration gradient exists for water molecules in the system. Water diffuses from the region of greater concentration of water (pure water outside) to the region of lesser concentration (salt solution inside).
As water enters the salt solution, the fluid level in the funnel rises. Gravity creates a hydrostatic pressure inside the osmometer. Eventually the pressure produced by the increasing weight of solution in the funnel pushes water molecules out as fast as they enter. The level in the funnel becomes stationary and the system is in equilibrium. The osmotic pressure of the solution is equivalent to the hydrostatic pressure necessary to prevent further net entry of water.
The concept of osmotic pressure is not without problems. A solution reveals an osmotic “pressure” only when it is separated from solvent by a selectively permeable membrane. It can be disconcerting to think of an isolated bottle of salt solution as having “pressure” much as compressed gas in a bottle (hydrostatic pressure) would have. Furthermore, the osmotic pressure is really the hydrostatic pressure that must be applied to a solution to keep it from gaining water if the solution were separated from pure water by a selectively permeable membrane. Consequently, biologists frequently use the term osmotic potential rather than osmotic pressure. However, since the term “osmotic pressure” is so firmly fixed in our vocabulary, it is necessary to understand the usage despite its potential confusion.
The concept of osmosis is very important in understanding how animals control their internal fluid and solute environment (see Homeostasis). For example, marine bony fishes maintain a solute concentration in their blood about one-third of that in seawater; they are hypoosmotic to seawater. If a fish swims into a river mouth and then up a freshwater stream, as salmon do, it would pass through a region where its blood solutes were equal in concentration to those in its environment (isosmotic), then enter fresh water, where its blood solutes were hyperosmotic to those in its environment. It must have physiological mechanisms to avoid net loss of water in the sea and gain of water in the river.
Mediated Transport
We have seen that the cell membrane
is an effective barrier to the free diffusion
of most molecules of biological
significance. Yet it is essential that such
materials enter and leave the cell.
Nutrients such as sugars and materials
for growth such as amino acids must
enter the cell, and the wastes of metabolism
must leave. Such molecules are
moved across the membrane by special
proteins called transporters or
permeases. Permeases form a small
passageway through the membrane,
enabling the solute molecule to cross
the phospholipid bilayer (Figure 3-18A).
Permeases are usually quite specific,
recognizing and transporting only a
limited group of chemical substances
or perhaps even a single substance.
At high concentrations of solute, mediated transport systems show a saturation effect. This means simply that the rate of influx reaches a plateau beyond which increasing the solute concentration has no further effect on influx rate (Figure 3-18B). This is evidence that the number of transporters available in the membrane is limited. When all transporters become occupied by solutes, the rate of transport is at a maximum and it cannot be increased. Simple diffusion shows no such limitation; the greater the difference in solute concentrations on the two sides of the membrane, the faster the influx.
Two distinctly different kinds of mediated transport mechanisms are recognized: (1) facilitated diffusion, in which the permease assists a molecule to diffuse through the membrane that it cannot otherwise penetrate, and (2) active transport, in which energy is supplied to the transporter system to transport molecules in the direction opposite a concentration gradient (Figure 3-19). Facilitated diffusion therefore differs from active transport in that it sponsors movement in a downhill direction (in the direction of the concentration gradient) only and requires no metabolic energy to drive the transport system.
In many animals facilitated diffusion
aids in the transport of glucose
(blood sugar) into body cells that oxidize
it as a principal energy source for
the synthesis of ATP. The concentration
of glucose is greater in the blood than
in the cells that consume it, favoring
inward diffusion, but glucose is a
water-soluble molecule that does not,
by itself, penetrate the membrane
rapidly enough to support the metabolism
of many cells; the carrier system
increases the inward flow of glucose.
In active transport, molecules are moved uphill against the forces of passive diffusion. Active transport always involves the expenditure of energy (from ATP) because materials are pumped against a concentration gradient. Among the most important active transport systems in all animals are those that maintain sodium and potassium ion gradients between cells and the surrounding extracellular fluid or external environment. Most animal cells require a high internal concentration of potassium ions for protein synthesis at the ribosome and for certain enzymatic functions. The potassium ion concentration may be 20 to 50 times greater inside the cell than outside. Sodium ions, on the other hand, may be 10 times more concentrated outside the cell than inside. Both of these ionic gradients are maintained by the active transport of potassium ions into and sodium ions out of the cell. In many cells the outward pumping of sodium is linked to the inward pumping of potassium; the same transporter molecule does both. As much as 10% to 40% of all the energy produced by the cell is consumed by the sodium-potassium exchange pump (Figure 3-19).
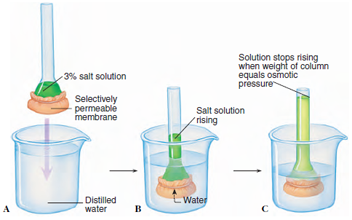 |
| Figure 3-17 Simple membrane osmometer. A, The end of a tube containing a salt solution is closed at one end by a selectively permeable membrane. The membrane is permeable to water but not to salt. B, When the tube is immersed in pure water, water molecules diffuse through the membrane into the tube. Water molecules are in higher concentration in the beaker because they are diluted inside the tube by salt ions. Because the salt cannot diffuse out through the membrane, the volume of fluid inside the tube increases, and the level rises. C, When the weight of the column of water inside the tube exerts a downward force (hydrostatic pressure) causing water molecules to leave through the membrane in equal number to those that enter, the volume of fluid inside the tube stops rising. At this point the hydrostatic pressure is equivalent to the osmotic pressure. |
Most cell membranes are selectively permeable, that is, permeable to water but variably permeable or impermeable to solutes. In free diffusion it is this selectiveness that regulates molecular traffic. As a rule, gases (such as oxygen and carbon dioxide), urea, and lipid-soluble solutes (such as hydrocarbons and alcohol) are the only solutes that can diffuse through biological membranes with any degree of freedom. Because many water-soluble molecules readily pass through membranes, such movements cannot be explained by simple diffusion. Sugars, as well as many electrolytes and macromolecules, are moved across membranes by carriermediated processes, which are described in the next section.
If we place a membrane between two unequal concentrations of solutes to which the membrane is impermeable, water flows through the membrane from the more dilute to the more concentrated solution. The water molecules move across the membrane down a concentration gradient from an area where the water molecules are more concentrated to an area on the other side of the membrane where they are less concentrated. This is osmosis.
We can demonstrate osmosis by a simple experiment in which we tie a selectively permeable membrane such as cellophane tightly over the end of a funnel. We fill the funnel with a salt solution and place it in a beaker of pure water so that the water levels inside and outside the funnel are equal. In a short time the water level in the glass tube of the funnel rises, indicating a net movement of water through the cellophane membrane into the salt solution (Figure 3-17).
Inside the funnel are salt molecules, as well as water molecules. In the beaker outside the funnel are only water molecules. Thus the concentration of water is less on the inside because some of the available space is occupied by the larger, nondiffusible salt molecules. A concentration gradient exists for water molecules in the system. Water diffuses from the region of greater concentration of water (pure water outside) to the region of lesser concentration (salt solution inside).
As water enters the salt solution, the fluid level in the funnel rises. Gravity creates a hydrostatic pressure inside the osmometer. Eventually the pressure produced by the increasing weight of solution in the funnel pushes water molecules out as fast as they enter. The level in the funnel becomes stationary and the system is in equilibrium. The osmotic pressure of the solution is equivalent to the hydrostatic pressure necessary to prevent further net entry of water.
The concept of osmotic pressure is not without problems. A solution reveals an osmotic “pressure” only when it is separated from solvent by a selectively permeable membrane. It can be disconcerting to think of an isolated bottle of salt solution as having “pressure” much as compressed gas in a bottle (hydrostatic pressure) would have. Furthermore, the osmotic pressure is really the hydrostatic pressure that must be applied to a solution to keep it from gaining water if the solution were separated from pure water by a selectively permeable membrane. Consequently, biologists frequently use the term osmotic potential rather than osmotic pressure. However, since the term “osmotic pressure” is so firmly fixed in our vocabulary, it is necessary to understand the usage despite its potential confusion.
The concept of osmosis is very important in understanding how animals control their internal fluid and solute environment (see Homeostasis). For example, marine bony fishes maintain a solute concentration in their blood about one-third of that in seawater; they are hypoosmotic to seawater. If a fish swims into a river mouth and then up a freshwater stream, as salmon do, it would pass through a region where its blood solutes were equal in concentration to those in its environment (isosmotic), then enter fresh water, where its blood solutes were hyperosmotic to those in its environment. It must have physiological mechanisms to avoid net loss of water in the sea and gain of water in the river.
Mediated Transport
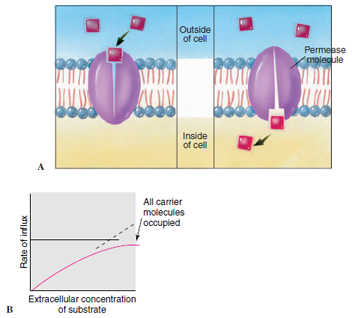 |
| Figure 3-18 Facilitated transport. A, The permease molecule binds with a molecule to be transported (substrate) on one side of the plasma membrane, changes shape, and releases the molecule on the other side. Facilitated transport takes place in the direction of a concentration gradient. B, Rate of transport increases with increasing substrate concentration until all permease molecules are occupied. |
At high concentrations of solute, mediated transport systems show a saturation effect. This means simply that the rate of influx reaches a plateau beyond which increasing the solute concentration has no further effect on influx rate (Figure 3-18B). This is evidence that the number of transporters available in the membrane is limited. When all transporters become occupied by solutes, the rate of transport is at a maximum and it cannot be increased. Simple diffusion shows no such limitation; the greater the difference in solute concentrations on the two sides of the membrane, the faster the influx.
Two distinctly different kinds of mediated transport mechanisms are recognized: (1) facilitated diffusion, in which the permease assists a molecule to diffuse through the membrane that it cannot otherwise penetrate, and (2) active transport, in which energy is supplied to the transporter system to transport molecules in the direction opposite a concentration gradient (Figure 3-19). Facilitated diffusion therefore differs from active transport in that it sponsors movement in a downhill direction (in the direction of the concentration gradient) only and requires no metabolic energy to drive the transport system.
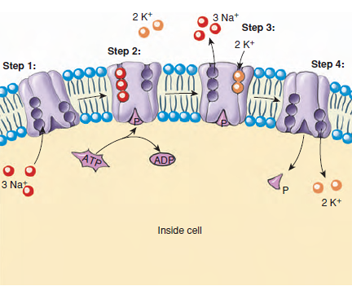 |
| Figure 3-19 Sodium-potassium pump, powered by bond energy of ATP, maintains the normal gradients of these ions across the cell membrane. The pump works by a series of conformational changes in the permease: Step 1. Three ions of Na+ bind to the interior end of the permease, producing a conformational (shape) change in the protein complex. +. The complex binds a molecule of ATP and cleaves it. Step 3. The binding of the phosphate group to the complex induces a second conformational change, passing the three Na+ ions across the membrane, where they are now positioned facing the exterior. This new conformation has a very low affinity for the Na+ ions, which dissociate and diffuse away, but it has a high affinity for K+ ions and binds two of them as soon as it is free of the Na+ ions. Step 4. Binding of the K+ ions leads to another conformational change in the complex, this time leading to dissociation of the bound phosphate. Freed of the phosphate, the complex reverts to its original conformation, with the two K+ ions exposed on the interior side of the membrane. This conformation has a low affinity for K+ ions so that they are now released, and the complex has the conformation it started with, having a high affinity for Na+ ions. |
In active transport, molecules are moved uphill against the forces of passive diffusion. Active transport always involves the expenditure of energy (from ATP) because materials are pumped against a concentration gradient. Among the most important active transport systems in all animals are those that maintain sodium and potassium ion gradients between cells and the surrounding extracellular fluid or external environment. Most animal cells require a high internal concentration of potassium ions for protein synthesis at the ribosome and for certain enzymatic functions. The potassium ion concentration may be 20 to 50 times greater inside the cell than outside. Sodium ions, on the other hand, may be 10 times more concentrated outside the cell than inside. Both of these ionic gradients are maintained by the active transport of potassium ions into and sodium ions out of the cell. In many cells the outward pumping of sodium is linked to the inward pumping of potassium; the same transporter molecule does both. As much as 10% to 40% of all the energy produced by the cell is consumed by the sodium-potassium exchange pump (Figure 3-19).
Support our developers




