Temperature Regulation in Endotherms
Temperature Regulation
in Endotherms
Most mammals have body temperatures between 36° and 38° C, somewhat lower than those of birds, which range between 40° and 42° C. Constant temperature is maintained by a delicate balance between heat production and heat loss—not a simple matter when these animals are alternating between periods of rest and bursts of activity.
Heat is produced by the animal’s metabolism. This includes oxidation of foods, basal cellular metabolism, and muscular contraction. Because much of an endotherm’s daily caloric intake is required to generate heat, especially in cold weather, the endotherm must eat more food than an ectotherm of the same size. Heat is lost by radiation, conduction, and convection (air movement) to a cooler environment and by evaporation of water (Figure 32-17). A bird or mammal can control both processes of heat production and heat loss within rather wide limits. If the animal becomes too cool, it can generate heat by increasing muscular activity (exercise or shivering) and by decreasing heat loss by increasing its insulation. If it becomes too warm, it decreases heat production and increases heat loss. We will examine these processes in the following examples.
Adaptations for Hot Environments
Despite the harsh conditions of deserts—intense heat during the day, cold at night, and scarcity of water, vegetation, and cover—many kinds of animals live there successfully. The smaller desert mammals are mostly fossorial (living mainly in the ground) or nocturnal (active at night). The lower temperature and higher humidity of burrows help to reduce water loss by evaporation. As explained earlier in this section, desert animals such as the kangaroo rat and the American desert ground squirrels can, if necessary, derive the water they need from their dry food, drinking no water at all. Such animals produce a highly concentrated urine and form almost completely dry feces.
Large desert ungulates (hooved mammals that chew their cud) obviously cannot escape desert heat by living in burrows. Animals such as camels and desert antelopes (gazelle, oryx, and eland) possess a number of adaptations for coping with heat and dehydration. Figure 32-18 shows those of the eland. Mechanisms for controlling water loss and preventing overheating are closely linked. The glossy, pallid color of fur reflects direct sunlight, and fur itself is an excellent insulation that resists heat. Heat is lost by convection and conduction from the underside of elands where the fur is very thin. Fat tissue, an essential food reserve, is concentrated in a single hump on the back, instead of being uniformly distributed under the skin where it would impair loss of heat by radiation. Elands avoid evaporative water loss—the only means an animal has for cooling itself when the environmental temperature is higher than that of the body—by permitting their body temperature to drop during the cool night and then to rise slowly during the day as the body stores heat. Only when the body temperature reaches 41° C must elands prevent further rise through evaporative cooling by sweating and panting. They conserve water by producing a concentrated urine and dry feces. Camels have all of these adaptations developed to a similar or even greater degree; they are perhaps the most perfectly adapted of all large desert mammals.
Adaptations for Cold Environments
In cold environments mammals and birds use two major mechanisms to maintain homeothermy: (1) decreased
conductance, reduction of heat loss by increasing the effectiveness of the insulation, and (2) increased heat production.
In all mammals living in the cold regions of the earth, fur thickness increases in winter, sometimes by as much as 50%. Thick underhair is the principal insulating layer, whereas the longer and more visible guard hair serves as protection against wear and for protective coloration. However, unlike the well-insulated trunk of the body, the body extremities (legs, tail, ears, nose) of arctic mammals are thinly insulated and exposed to rapid cooling. To prevent these parts from becoming major avenues of heat loss, they are allowed to cool to low temperatures, often approaching the freezing point. The heat in the warm arterial blood is not lost from the body, however. Instead, a countercurrent heat exchange between the outgoing warm blood and the returning cold blood prevents heat loss. Arterial blood in the leg of an arctic mammal or bird passes in close contact with a network of small veins. Because arterial blood flow is opposite to that of returning venous blood, heat is exchanged very efficiently from artery to veins. By the time the arterial blood reaches the foot it has transferred nearly all of its heat to the veins returning blood to the body core (Figure 32- 19). Thus little heat is lost from poorly insulated distal regions of the leg to the surrounding cold air. Countercurrent heat exchangers in appendages also are common in aquatic mammals such as seals and whales, which have thinly insulated flippers and flukes that would be avenues of excessive heat loss in the absence of this heatsalvaging arrangement.
A consequence of peripheral heat exchange is that legs and feet of mammals and birds living in cold environments must function at low temperatures. Temperatures of the feet of arctic foxes and barren-ground caribou are just above the freezing point; in fact, the temperature may be below 0° C in footpads and hooves. To keep feet supple and flexible at such low temperatures, fats in the extremities have very low melting points, perhaps 30° C lower than ordinary body fats. In severely cold conditions all mammals can produce more heat by aug-mented muscular activity through exercise or shivering. We are all familiar with the effectiveness of both activities. A person can increase heat production as much as 18-fold by violent shivering when maximally stressed by cold. Another source of heat is increased oxidation of foods, especially from stores of brown fat (brown fat is described on). This mechanism is called nonshivering thermogenesis.
Small mammals the size of lemmings, voles, and mice meet the challenge of cold environments in a different way. Small mammals are not as well insulated as large mammals because thickness of fur is limited by the need to maintain mobility. Consequently these forms exploit the excellent insulating qualities of snow by living under it in runways on the forest floor, where incidentally, their food also is located. In this subnivean environment the temperature seldom drops below −5° C even though the air temperature above may fall to −50° C. Snow insulation decreases thermal conductance from small mammals just as thick pelage does for large mammals. Living beneath the snow is really a type of avoidance response to cold.
Most mammals have body temperatures between 36° and 38° C, somewhat lower than those of birds, which range between 40° and 42° C. Constant temperature is maintained by a delicate balance between heat production and heat loss—not a simple matter when these animals are alternating between periods of rest and bursts of activity.
Heat is produced by the animal’s metabolism. This includes oxidation of foods, basal cellular metabolism, and muscular contraction. Because much of an endotherm’s daily caloric intake is required to generate heat, especially in cold weather, the endotherm must eat more food than an ectotherm of the same size. Heat is lost by radiation, conduction, and convection (air movement) to a cooler environment and by evaporation of water (Figure 32-17). A bird or mammal can control both processes of heat production and heat loss within rather wide limits. If the animal becomes too cool, it can generate heat by increasing muscular activity (exercise or shivering) and by decreasing heat loss by increasing its insulation. If it becomes too warm, it decreases heat production and increases heat loss. We will examine these processes in the following examples.
Adaptations for Hot Environments
Despite the harsh conditions of deserts—intense heat during the day, cold at night, and scarcity of water, vegetation, and cover—many kinds of animals live there successfully. The smaller desert mammals are mostly fossorial (living mainly in the ground) or nocturnal (active at night). The lower temperature and higher humidity of burrows help to reduce water loss by evaporation. As explained earlier in this section, desert animals such as the kangaroo rat and the American desert ground squirrels can, if necessary, derive the water they need from their dry food, drinking no water at all. Such animals produce a highly concentrated urine and form almost completely dry feces.
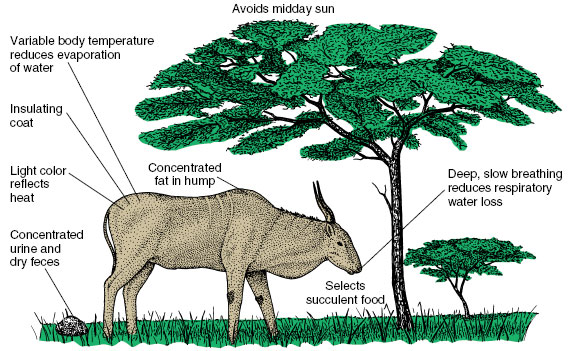 |
| Figure 32-18 Physiological and behavioral adaptations of the common eland for regulating temperature in the hot, arid savanna of central Africa. |
Large desert ungulates (hooved mammals that chew their cud) obviously cannot escape desert heat by living in burrows. Animals such as camels and desert antelopes (gazelle, oryx, and eland) possess a number of adaptations for coping with heat and dehydration. Figure 32-18 shows those of the eland. Mechanisms for controlling water loss and preventing overheating are closely linked. The glossy, pallid color of fur reflects direct sunlight, and fur itself is an excellent insulation that resists heat. Heat is lost by convection and conduction from the underside of elands where the fur is very thin. Fat tissue, an essential food reserve, is concentrated in a single hump on the back, instead of being uniformly distributed under the skin where it would impair loss of heat by radiation. Elands avoid evaporative water loss—the only means an animal has for cooling itself when the environmental temperature is higher than that of the body—by permitting their body temperature to drop during the cool night and then to rise slowly during the day as the body stores heat. Only when the body temperature reaches 41° C must elands prevent further rise through evaporative cooling by sweating and panting. They conserve water by producing a concentrated urine and dry feces. Camels have all of these adaptations developed to a similar or even greater degree; they are perhaps the most perfectly adapted of all large desert mammals.
Adaptations for Cold Environments
In cold environments mammals and birds use two major mechanisms to maintain homeothermy: (1) decreased
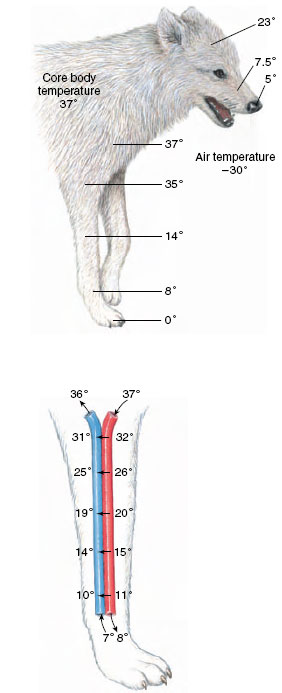 |
| Figure 32-19 Countercurrent heat exchange in the leg of an arctic wolf. The upper diagram shows how the extremities cool when the animal is exposed to low air temperatures. The lower diagram depicts a portion of the front leg artery and vein, showing how heat is exchanged between arterial and venous blood. Heat is shunted back into the body and conserved. |
conductance, reduction of heat loss by increasing the effectiveness of the insulation, and (2) increased heat production.
In all mammals living in the cold regions of the earth, fur thickness increases in winter, sometimes by as much as 50%. Thick underhair is the principal insulating layer, whereas the longer and more visible guard hair serves as protection against wear and for protective coloration. However, unlike the well-insulated trunk of the body, the body extremities (legs, tail, ears, nose) of arctic mammals are thinly insulated and exposed to rapid cooling. To prevent these parts from becoming major avenues of heat loss, they are allowed to cool to low temperatures, often approaching the freezing point. The heat in the warm arterial blood is not lost from the body, however. Instead, a countercurrent heat exchange between the outgoing warm blood and the returning cold blood prevents heat loss. Arterial blood in the leg of an arctic mammal or bird passes in close contact with a network of small veins. Because arterial blood flow is opposite to that of returning venous blood, heat is exchanged very efficiently from artery to veins. By the time the arterial blood reaches the foot it has transferred nearly all of its heat to the veins returning blood to the body core (Figure 32- 19). Thus little heat is lost from poorly insulated distal regions of the leg to the surrounding cold air. Countercurrent heat exchangers in appendages also are common in aquatic mammals such as seals and whales, which have thinly insulated flippers and flukes that would be avenues of excessive heat loss in the absence of this heatsalvaging arrangement.
A consequence of peripheral heat exchange is that legs and feet of mammals and birds living in cold environments must function at low temperatures. Temperatures of the feet of arctic foxes and barren-ground caribou are just above the freezing point; in fact, the temperature may be below 0° C in footpads and hooves. To keep feet supple and flexible at such low temperatures, fats in the extremities have very low melting points, perhaps 30° C lower than ordinary body fats. In severely cold conditions all mammals can produce more heat by aug-mented muscular activity through exercise or shivering. We are all familiar with the effectiveness of both activities. A person can increase heat production as much as 18-fold by violent shivering when maximally stressed by cold. Another source of heat is increased oxidation of foods, especially from stores of brown fat (brown fat is described on). This mechanism is called nonshivering thermogenesis.
Small mammals the size of lemmings, voles, and mice meet the challenge of cold environments in a different way. Small mammals are not as well insulated as large mammals because thickness of fur is limited by the need to maintain mobility. Consequently these forms exploit the excellent insulating qualities of snow by living under it in runways on the forest floor, where incidentally, their food also is located. In this subnivean environment the temperature seldom drops below −5° C even though the air temperature above may fall to −50° C. Snow insulation decreases thermal conductance from small mammals just as thick pelage does for large mammals. Living beneath the snow is really a type of avoidance response to cold.
Support our developers




