Water Excretion
Water Excretion
The kidney closely regulates the osmotic pressure of the blood. When fluid intake is high, the kidney excretes a dilute urine, saving salts and excreting water. When fluid intake is low, the kidney conserves water by forming a concentrated urine. A dehydrated person can concentrate urine to approximately four times blood osmotic concentration. This important ability to concentrate urine enables us to excrete wastes with minimal loss of water.
The capacity of the kidney of mammals and some birds to produce a concentrated urine involves an interaction between the loop of Henle and the collecting ducts. This interplay results in the formation of an osmotic gradient in the kidney, as shown in Figure 32-14. In the cortex, the interstitial fluid is isosmotic with the blood, but deep in the medulla the osmotic concentration is 4 times greater than that of the blood (in rodents and desert mammals that can produce highly concentrated urine the osmotic gradient is much greater than in humans). The high osmotic concentrations in the medulla are produced by an exchange of ions in the loop of Henle by countercurrent multiplication. “Countercurrent” refers to the opposite directions of fluid movement in the two limbs of the loop of Henle: down in the descending limb and up the ascending limb. “Multiplication” describes the increasing osmotic concentration in the medulla resulting from ion exchange between the two limbs of the loop.
The functional characteristics of this system are as follows. The descending limb of the loop of Henle is permeable to water but impermeable to solutes. The ascending limb is relatively impermeable to both water and solutes. Sodium chloride is actively transported out of the thick portion of the ascending limb and into the surrounding tissue fluid (Figure 32-14). As the interstitium surrounding the loop becomes more concentrated with solute, water is withdrawn from the descending limb by osmosis. The tubular fluid in the base of the loop, now more concentrated, moves up the ascending limb, where still more sodium chloride is pumped out. In this way the effect of active ion transport in the ascending limb is multiplied as more water is withdrawn from the descending limb and more concentrated fluid is presented to the ascending limb ion pump (Figures 32-14 and 32-15).
Final adjustment of urine concentration occurs not in the loops of Henle but in the collecting ducts. Formative urine that enters the distal tubule from the loop of Henle is dilute (because of active salt withdrawal) and is diluted still more by active reabsorption of more sodium chloride in the distal tubule. The formative urine, low in solutes but carrying urea, now flows down into the collecting duct. Because of the high concentration of solutes surrounding the collecting duct, water is withdrawn from the urine. As the urine becomes more concentrated, urea also diffuses out and adds to the high osmotic pressure in the kidney medulla (Figure 32-15).
The amount of water reabsorbed and the final concentration of the urine depend on the permeability of the walls of the distal convoluted tubule and the collecting duct. This is controlled by the antidiuretic hormone (ADH, or vasopressin), which is released by the posterior pituitary gland (neurohypophysis). In turn, special receptors in the brain that constantly sense the osmotic pressure of the body fluids govern the release of this hormone. When the blood osmotic pressure increases, as during dehydration, the pituitary gland releases more ADH. ADH increases the permeability of the collecting duct, probably by expanding the size of pores in the walls of the duct. Then, as the fluid in the collecting duct passes through the hyperosmotic region of the kidney medulla, water diffuses through the pores into the surrounding interstitial fluid and is carried away by the blood circulation. The urine loses water and becomes more concentrated. Given this sequence of events for dehydration, it is not difficult to anticipate how the system responds to overhydration: the pituitary stops releasing ADH, the pores in the collecting duct walls close, and a large volume of dilute urine is excreted.
The varying ability of different mammals to form a concentrated urine correlates closely with length of the loops of Henle. The beaver, which has no need to conserve water in its aquatic environment, has short loops and can concentrate its urine only to about twice the osmolarity of its blood plasma. Humans, with relatively longer loops, can concentrate urine 4.2 times that of the blood. As we would anticipate, desert mammals have much greater urine concentrating powers. A camel can produce a urine 8 times the plasma concentration, a gerbil 14 times, and an Australian hopping mouse 22 times. In this creature, the greatest urine concentrator of all, the loops of Henle extend to the tip of a long renal papilla that pushes out into the mouth of the ureter.
The kidney closely regulates the osmotic pressure of the blood. When fluid intake is high, the kidney excretes a dilute urine, saving salts and excreting water. When fluid intake is low, the kidney conserves water by forming a concentrated urine. A dehydrated person can concentrate urine to approximately four times blood osmotic concentration. This important ability to concentrate urine enables us to excrete wastes with minimal loss of water.
The capacity of the kidney of mammals and some birds to produce a concentrated urine involves an interaction between the loop of Henle and the collecting ducts. This interplay results in the formation of an osmotic gradient in the kidney, as shown in Figure 32-14. In the cortex, the interstitial fluid is isosmotic with the blood, but deep in the medulla the osmotic concentration is 4 times greater than that of the blood (in rodents and desert mammals that can produce highly concentrated urine the osmotic gradient is much greater than in humans). The high osmotic concentrations in the medulla are produced by an exchange of ions in the loop of Henle by countercurrent multiplication. “Countercurrent” refers to the opposite directions of fluid movement in the two limbs of the loop of Henle: down in the descending limb and up the ascending limb. “Multiplication” describes the increasing osmotic concentration in the medulla resulting from ion exchange between the two limbs of the loop.
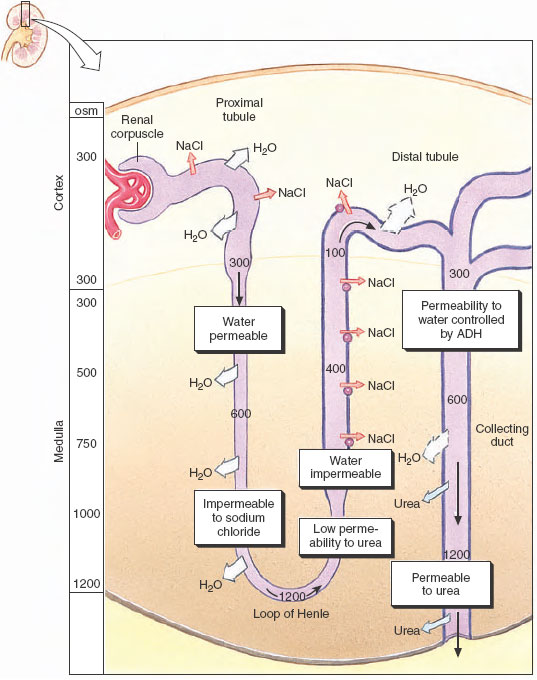 |
| Figure 32-14 Mechanism of urine concentration in mammals. Sodium and chloride are pumped from the ascending limb of the loop of Henle, and water is withdrawn passively from the descending limb, which is impermeable to sodium chloride. Sodium chloride and urea reabsorbed from the collecting duct raise the osmotic concentration in the kidney medulla, creating an osmotic gradient for the controlled reabsorption of water from the collecting duct. |
The functional characteristics of this system are as follows. The descending limb of the loop of Henle is permeable to water but impermeable to solutes. The ascending limb is relatively impermeable to both water and solutes. Sodium chloride is actively transported out of the thick portion of the ascending limb and into the surrounding tissue fluid (Figure 32-14). As the interstitium surrounding the loop becomes more concentrated with solute, water is withdrawn from the descending limb by osmosis. The tubular fluid in the base of the loop, now more concentrated, moves up the ascending limb, where still more sodium chloride is pumped out. In this way the effect of active ion transport in the ascending limb is multiplied as more water is withdrawn from the descending limb and more concentrated fluid is presented to the ascending limb ion pump (Figures 32-14 and 32-15).
Final adjustment of urine concentration occurs not in the loops of Henle but in the collecting ducts. Formative urine that enters the distal tubule from the loop of Henle is dilute (because of active salt withdrawal) and is diluted still more by active reabsorption of more sodium chloride in the distal tubule. The formative urine, low in solutes but carrying urea, now flows down into the collecting duct. Because of the high concentration of solutes surrounding the collecting duct, water is withdrawn from the urine. As the urine becomes more concentrated, urea also diffuses out and adds to the high osmotic pressure in the kidney medulla (Figure 32-15).
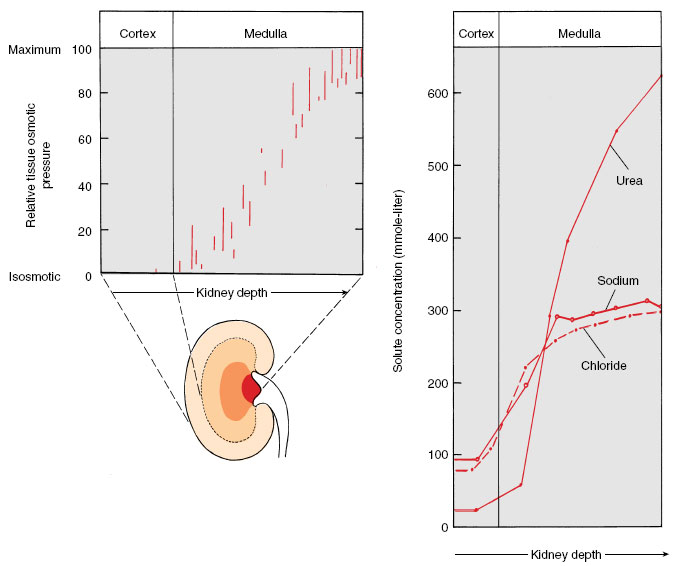 |
| Figure 32-15 Osmotic concentration of tissue fluid in the mammalian kidney. Tissue fluid is isosmotic in the kidney cortex (to left in diagram) but osmotic concentration increases continuously through the medulla, reaching a maximum at the papilla where the urine drains into the ureter. |
The amount of water reabsorbed and the final concentration of the urine depend on the permeability of the walls of the distal convoluted tubule and the collecting duct. This is controlled by the antidiuretic hormone (ADH, or vasopressin), which is released by the posterior pituitary gland (neurohypophysis). In turn, special receptors in the brain that constantly sense the osmotic pressure of the body fluids govern the release of this hormone. When the blood osmotic pressure increases, as during dehydration, the pituitary gland releases more ADH. ADH increases the permeability of the collecting duct, probably by expanding the size of pores in the walls of the duct. Then, as the fluid in the collecting duct passes through the hyperosmotic region of the kidney medulla, water diffuses through the pores into the surrounding interstitial fluid and is carried away by the blood circulation. The urine loses water and becomes more concentrated. Given this sequence of events for dehydration, it is not difficult to anticipate how the system responds to overhydration: the pituitary stops releasing ADH, the pores in the collecting duct walls close, and a large volume of dilute urine is excreted.
The varying ability of different mammals to form a concentrated urine correlates closely with length of the loops of Henle. The beaver, which has no need to conserve water in its aquatic environment, has short loops and can concentrate its urine only to about twice the osmolarity of its blood plasma. Humans, with relatively longer loops, can concentrate urine 4.2 times that of the blood. As we would anticipate, desert mammals have much greater urine concentrating powers. A camel can produce a urine 8 times the plasma concentration, a gerbil 14 times, and an Australian hopping mouse 22 times. In this creature, the greatest urine concentrator of all, the loops of Henle extend to the tip of a long renal papilla that pushes out into the mouth of the ureter.
Support our developers




