Cell-Specific Expression of Monoterpenoid Indole Alkaloid Biosynthetic Genes
A detailed study of the expression of several genes of vindoline biosynthesis in developing C. roseus leaves using in situ hybridization and immunolocalization provides the first clear insight into the spatial distribution of monoterpenoid indole alkaloid biosynthesis (St-Pierre et al., 1999). in situ hybridization of two genes, tdc and str1, occurring early in the vindoline biosynthetic pathway and two genes, d4h and dat, occurring late in the vindoline biosynthesis revealed that multiple cell types are involved. C. roseus produces more than 180 monoterpenoid indole alkaloids all of which are derived from the central intermediate 3α(S)- strictosidine. The enzymes tryptophan decarboxylase and strictosidine synthase lead to this central intermediate and are, therefore, involved in the biosynthesis of all of the C. roseus alkaloids. The transcripts of tdc and str1 were found in the epidermis of developing leaves (Fig. 10.4A–D, K, and L) (Irmler et al., 2000; St-Pierre et al., 1999). In contrast, transcripts of the vindoline-specific biosynthetic genes d4h and dat localized to laticifer and idioblasts of developing leaves (Fig. 10.4E–H, M, and N) (Irmler et al., 2000; St-Pierre et al., 1999). In addition, transcript of the secologanin biosynthetic enzyme
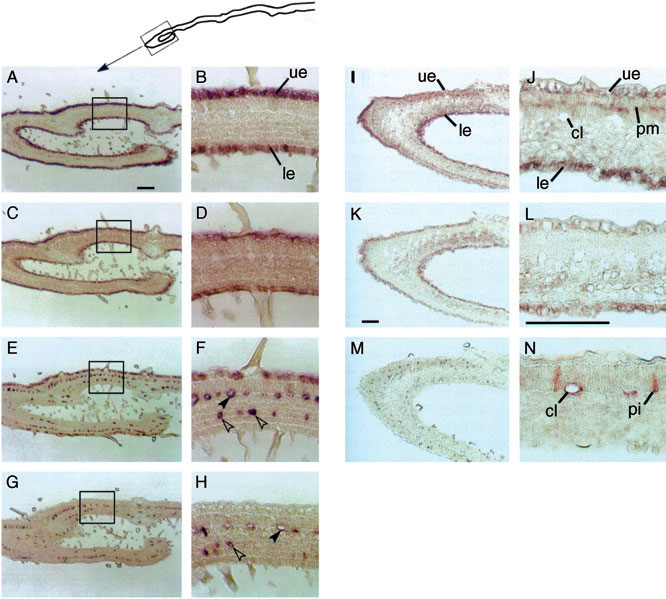 |
| FIGURE 10.4 Localization of cyp72a1, tdc, str1, d4h, and dat transcripts in developing C. roseus leaves (Irmler et al., 2000; St-Pierre et al., 1999). Panels (A, B, K, L) tdc; panels (C, D) str1; panels (E, F) d4h; panels (G, H, M, N) dat; panels (I, J) cyp72a1. cl, cross-connecting laticifer cells; le, lower epidermis; pi, idioblast cells associated with palisade mesophyll cells; pm, palisade mesophyll cells; si, idioblast cells associated with spongy mesophyll cells; ue, upper epidermis; tdc, tryptophan decarboxylase; str1, strictosidine synthase; cyp72a1, secologanin synthase; d4h, desacetoxyvindoline 4-hydroxlyase; dat, deacetylvindoline 4-O-acetyltransferase. Bar shown in panels (A, K), 100 µm; in panel (L), 50 µm. Solid arrows in panels (F, H), laticifer cells; open arrows, idioblast cells. |
Transcript of tdc and str1 are also found in protoderm and cortical cells around the apical meristem of root tips (St-Pierre et al., 1999). Likewise, the transcript of a gene involved in the biosynthesis of the root-specific monoterpenoid indole alkaloid minovincinine was detected in root tissue. RNA in situ hybridization studies located minovincinine 19-hydroxy-O-acetyltransferase gene expression within the cortex and epidermis of tissues near the root tip (Laflamme et al., 2001).
Multicellular compartmentation of alkaloid biosynthesis should be a central consideration in the metabolic engineering of alkaloid pathways. Promoters should be chosen that would direct transgene expression to the cell types in which the appropriate biosynthetic intermediates are expected to occur or in which biosynthetic genes are expressed.
Support our developers




