Cytoplasmic Specification
Cytoplasmic Specification
A fertilized egg contains cytoplasmic
components that are unequally distributed
within the egg. These different
cytoplasmic components are thought
to contain morphogenetic determinants
that control commitment of the
cell to a particular cell type. These
morphogenetic determinants are partitioned
among different blastomeres as
a result of cleavage, and the developmental
fate of each cell becomes specified
by the type of cytoplasm it
acquires during development.
This process is especially striking (and easily visualized) in some tunicate species in which the fertilized egg contains as many as five differently colored types of cytoplasm (Figure 8-10). These differently pigmented cytoplasms are segregated into different blastomeres which then proceed to form distinct tissues or organs. For example, yellow cytoplasm gives rise to muscle cells while gray equatorial cytoplasm produces the notochord and neural tube. Clear cytoplasm produces the larval epidermis and gray vegetal cytoplasm gives rise to the gut.
Cytoplasmic specification is less important in vertebrate embryos, but it is seen to some extent. For example, in Spemann’s experiment, normal development cannot occur in the absence of gray-crescent cytoplasm (see Figure 8- 13B). Cells that receive gray-crescent cytoplasm form the dorsal lip of the blastopore, and without this cytoplasm the dorsal lip does not form and gastrulation cannot occur, leading to abnormal development.
Another characteristic of this type of specification is that cell fate is determined without reference to neighboring cells. When a particular blastomere is isolated from the rest of the embryo, it still forms its characteristic structure (Figure 8-14B). In the absence of a particular blastomere, the animal lacks just those structures normally formed by that blastomere. This pattern is called mosaic development, since the embryo seems to be a mosaic of selfdifferentiating parts. Mosaic development is characteristic of most protostomes (see Figure 8-9).
In many animals, the fate of a cell depends on its interactions with neighboring cells, rather than on what piece of cytoplasm it acquired during cleavage. In these embryos, at least early in development, each cell is able to produce an entire embryo if separated from the other cells (see Figure 8-14A). In other words, an early blastomere originally has the ability to follow more than one path of differentiation, but its interaction with other cells restricts its fate. If a blastomere is removed from an early embryo, the remaining blastomeres can alter their normal fates so as to compensate for the missing blastomere and produce a complete organism. This adaptability is termed regulative development. Regulative development occurs in most deuterostomes (excluding tunicates) (see Figure 8-9).
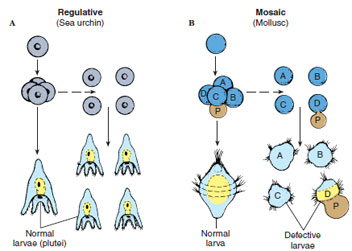 |
| Figure 8-14 Regulative and mosaic cleavage. A, Regulative cleavage. Each of the early blastomeres (such as that of a sea urchin) when separated from the others develops into a small pluteus larva. B, Mosaic cleavage. In the mollusc, when the blastomeres are separated, each gives rise to only a part of an embryo. The larger size of one of the defective larvae is the result of the formation of a polar lobe (P) composed of clear cytoplasm of the vegetal pole, which this blastomere alone receives. |
This process is especially striking (and easily visualized) in some tunicate species in which the fertilized egg contains as many as five differently colored types of cytoplasm (Figure 8-10). These differently pigmented cytoplasms are segregated into different blastomeres which then proceed to form distinct tissues or organs. For example, yellow cytoplasm gives rise to muscle cells while gray equatorial cytoplasm produces the notochord and neural tube. Clear cytoplasm produces the larval epidermis and gray vegetal cytoplasm gives rise to the gut.
Cytoplasmic specification is less important in vertebrate embryos, but it is seen to some extent. For example, in Spemann’s experiment, normal development cannot occur in the absence of gray-crescent cytoplasm (see Figure 8- 13B). Cells that receive gray-crescent cytoplasm form the dorsal lip of the blastopore, and without this cytoplasm the dorsal lip does not form and gastrulation cannot occur, leading to abnormal development.
Another characteristic of this type of specification is that cell fate is determined without reference to neighboring cells. When a particular blastomere is isolated from the rest of the embryo, it still forms its characteristic structure (Figure 8-14B). In the absence of a particular blastomere, the animal lacks just those structures normally formed by that blastomere. This pattern is called mosaic development, since the embryo seems to be a mosaic of selfdifferentiating parts. Mosaic development is characteristic of most protostomes (see Figure 8-9).
In many animals, the fate of a cell depends on its interactions with neighboring cells, rather than on what piece of cytoplasm it acquired during cleavage. In these embryos, at least early in development, each cell is able to produce an entire embryo if separated from the other cells (see Figure 8-14A). In other words, an early blastomere originally has the ability to follow more than one path of differentiation, but its interaction with other cells restricts its fate. If a blastomere is removed from an early embryo, the remaining blastomeres can alter their normal fates so as to compensate for the missing blastomere and produce a complete organism. This adaptability is termed regulative development. Regulative development occurs in most deuterostomes (excluding tunicates) (see Figure 8-9).
Support our developers




