Gastrulation and the Formation of Germ Layers
Gastrulation and the
Formation of Germ
Layers
Gastrulation involves extensive and highly integrated cell and tissue movements, resulting in dramatic rearrangement of cells of the blastula. Gastrulation converts the spherical blastula into a more complex configuration of three germ layers. At the end of gastrulation, the ectoderm covers the embryo, and the mesoderm and endoderm have been brought inside. As a result, cells have new positions and new neighbors, and the interaction of these cells and tissues will generate the embryonic body plan. Patterns of gastrulation vary enormously between different groups of animals, and these differences depend very much on the amount and distribution of yolk. As with cleavage, yolk impedes gastrulation. Thus, gastrulation is relatively simple in most non-yolky embryos, but it is more complex in embryos developing from yolk-laden eggs.
In sea stars, gastrulation begins when the entire vegetal area of the blastula flattens to form the vegetal plate. This event is followed by a process called invagination, in which the vegetal plate (a sheet of epithelial tissue) bends inward and extends about one-third of the way into the blastocoel, forming a new internal cavity, the archenteron (Figure 8-11A). The archenteron is the primitive gut and its opening to the outside is called the blastopore. In sea stars and other members of the Deuterostomia (“mouth second”), the blastopore becomes the anus, while the mouth forms secondarily (see Figure 8-9). The archenteron continues to elongate toward the animal pole and its anterior end expands into two pouchlike coelomic vesicles, which pinch off to form left and right coelomic compartments (Figure 8-11A).
The gastrula is now an embryo of three germ layers. The outer layer is ectoderm; it will give rise to the epithelium of the body surface and to the nervous system. The inner layer that forms the archenteron is endoderm; it will give rise to the epithelial lining of the digestive tube. The outpocketing of the archenteron is the origin of mesoderm. This third germ layer will form the muscular system, reproductive system, peritoneum (lining of the coelomic compartments), and the calcareous plates of the sea star’s endoskeleton. The mesoderm is also the origin of the water vascular system of sea stars, a system unique to echinoderms.
Gastrulation in nemertean worms (see Figure 8-11C) resembles gastrulation in sea stars, in that the archenteron is formed by invagination. However, in nemerteans and other members of the Protostomia (“mouth first”), the blastopore becomes the mouth and the anus forms secondarily (see Figure 8-9). In addition, the mesoderm forms differently in protostomes and deuterostomes. In protostomes, cells destined to become mesoderm arise ventrally at the lip of the blastopore and proliferate between the walls of the archenteron (endoderm) and outer body wall (ectoderm). Meticulous cell lineage studies by early embryologists established that in many protostomes (for example, flatworms, annelids, and molluscs) these mesodermal precursors arise from a single large blastomere at the 29- to 64-cell stage embryo called the 4d cell (see Figure 10-13). In most nemerteans, the precise origin of the mesoderm is not yet known; in some it is probably the 4d cell, but in others it apparently derives from an earlier blastomere.
In frogs, deuterostomes with radial cleavage (see Figure 8-7B), the morphogenetic movements of gastrulation are greatly influenced by the mass of inert yolk in the vegetal half of the embryo. Cleavage divisions are slowed in this half so that the resulting blastula consists of many small cells in the animal half and a few large cells in the vegetal half (see Figure 8-11B). Gastrulation in amphibians begins when cells located at the future dorsal side of the embryo invaginate to form a slitlike blastopore. Thus, as in sea stars, invagination initiates archenteron formation, but amphibian gastrulation begins in the marginal zone of the blastula, where animal and vegetal hemispheres come together, and where there is less yolk than in the vegetal region. Gastrulation progresses as the sheets of cells in the marginal zone turn inward over the blastopore lip and move inside the gastrula to form mesoderm and endoderm (Figure 8-11B). The three germ layers now formed are the pri-mary structural layers that play crucial roles in further differentiation of the embryo.
In bird and reptile embryos (see
Figure 8-11D), gastrulation begins
with a thickening of the blastoderm at
the caudal end of the embryo that
migrates forward to form the primitive
streak (Figure 8-12). The primitive
streak becomes the anteroposterior
axis of the embryo and the center
of early growth. The primitive streak
is homologous to the blastopore of
frog embryos, but in the chick it does
not open into the gut cavity because
of the obstructing mass of yolk. The
blastoderm consists of two layers (epiblast
and hypoblast) with a blastocoel
between them. Cells of the epiblast
move as a sheet toward the primitive
streak, then roll over the edge and
migrate as individual cells into the
blastocoel. These migrating cells separate
into two streams. One stream of
cells moves deeper (displacing the
hypoblast along the midline) and
forms endoderm. The other stream
moves between the epiblast and
hypoblast to form mesoderm. Cells on
the surface of the embryo compose
the ectoderm. The embryo now has
three germ layers, at this point arranged
as sheetlike layers with ectoderm
on top and endoderm at the
bottom. This arrangement changes,
however, when all three germ layers
lift from the underlying yolk (Figure
8-12), then fold under to form a
three-layered embryo that is pinched
off from the yolk except for a stalk
attachment to the yolk at midbody.
Gastrulation in mammals is remarkably similar to gastrulation in reptiles and birds (see Figure 8-11E). Gastrulation movements in the inner cell mass produce a primitive streak. Epiblast cells move medially through the primitive streak into the blastocoel, and individual cells then migrate laterally through the blastocoel to form mesoderm and endoderm. Endoderm cells (derived from the hypoblast) form a yolk sac devoid of yolk (since the mammalian embryos derive nutrients directly from the mother via the placenta).
Amphibians, reptiles, and birds, which have moderate to large amounts of yolk concentrated in the vegetal region of the egg, have evolved derived gastrulation patterns in which the yolk does not participate in gastrulation. Yolk is an impediment to gastrulation and consequently the gastrulation process occurs around (amphibians) or on top (reptiles and birds) of the vegetal yolk. Mammalian eggs are isolecithal, and thus one might expect them to have a gastrulation pattern similar to that of sea stars. Instead they have a pattern more suited to telolecithal eggs. The best explanation for this feature of mammalian egg development is common ancestry with birds and reptiles. Reptiles, birds, and mammals share a common ancestor whose eggs were telolecithal. Thus, all three groups inherited their gastrulation patterns from this common ancestor, and mammals subsequently evolved isolecithal eggs but retained the telolecithal pattern.
In Cnidaria and Ctenophora, only two germ layers are formed, endoderm and ectoderm. These animals are diploblastic. In all other metazoa, the mesoderm also appears, either from pouches of the archenteron or from other cells associated with endoderm formation. This three-layered condition is called triploblastic.
Gastrulation involves extensive and highly integrated cell and tissue movements, resulting in dramatic rearrangement of cells of the blastula. Gastrulation converts the spherical blastula into a more complex configuration of three germ layers. At the end of gastrulation, the ectoderm covers the embryo, and the mesoderm and endoderm have been brought inside. As a result, cells have new positions and new neighbors, and the interaction of these cells and tissues will generate the embryonic body plan. Patterns of gastrulation vary enormously between different groups of animals, and these differences depend very much on the amount and distribution of yolk. As with cleavage, yolk impedes gastrulation. Thus, gastrulation is relatively simple in most non-yolky embryos, but it is more complex in embryos developing from yolk-laden eggs.
In sea stars, gastrulation begins when the entire vegetal area of the blastula flattens to form the vegetal plate. This event is followed by a process called invagination, in which the vegetal plate (a sheet of epithelial tissue) bends inward and extends about one-third of the way into the blastocoel, forming a new internal cavity, the archenteron (Figure 8-11A). The archenteron is the primitive gut and its opening to the outside is called the blastopore. In sea stars and other members of the Deuterostomia (“mouth second”), the blastopore becomes the anus, while the mouth forms secondarily (see Figure 8-9). The archenteron continues to elongate toward the animal pole and its anterior end expands into two pouchlike coelomic vesicles, which pinch off to form left and right coelomic compartments (Figure 8-11A).
The gastrula is now an embryo of three germ layers. The outer layer is ectoderm; it will give rise to the epithelium of the body surface and to the nervous system. The inner layer that forms the archenteron is endoderm; it will give rise to the epithelial lining of the digestive tube. The outpocketing of the archenteron is the origin of mesoderm. This third germ layer will form the muscular system, reproductive system, peritoneum (lining of the coelomic compartments), and the calcareous plates of the sea star’s endoskeleton. The mesoderm is also the origin of the water vascular system of sea stars, a system unique to echinoderms.
Gastrulation in nemertean worms (see Figure 8-11C) resembles gastrulation in sea stars, in that the archenteron is formed by invagination. However, in nemerteans and other members of the Protostomia (“mouth first”), the blastopore becomes the mouth and the anus forms secondarily (see Figure 8-9). In addition, the mesoderm forms differently in protostomes and deuterostomes. In protostomes, cells destined to become mesoderm arise ventrally at the lip of the blastopore and proliferate between the walls of the archenteron (endoderm) and outer body wall (ectoderm). Meticulous cell lineage studies by early embryologists established that in many protostomes (for example, flatworms, annelids, and molluscs) these mesodermal precursors arise from a single large blastomere at the 29- to 64-cell stage embryo called the 4d cell (see Figure 10-13). In most nemerteans, the precise origin of the mesoderm is not yet known; in some it is probably the 4d cell, but in others it apparently derives from an earlier blastomere.
In frogs, deuterostomes with radial cleavage (see Figure 8-7B), the morphogenetic movements of gastrulation are greatly influenced by the mass of inert yolk in the vegetal half of the embryo. Cleavage divisions are slowed in this half so that the resulting blastula consists of many small cells in the animal half and a few large cells in the vegetal half (see Figure 8-11B). Gastrulation in amphibians begins when cells located at the future dorsal side of the embryo invaginate to form a slitlike blastopore. Thus, as in sea stars, invagination initiates archenteron formation, but amphibian gastrulation begins in the marginal zone of the blastula, where animal and vegetal hemispheres come together, and where there is less yolk than in the vegetal region. Gastrulation progresses as the sheets of cells in the marginal zone turn inward over the blastopore lip and move inside the gastrula to form mesoderm and endoderm (Figure 8-11B). The three germ layers now formed are the pri-mary structural layers that play crucial roles in further differentiation of the embryo.
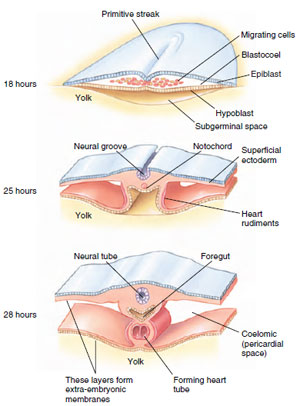 |
| Figure 8-12 Gastrulation in the chick. Transverse sections through the heart-forming region of the chick show development at 18, 25, and 28 hours of incubation. |
Gastrulation in mammals is remarkably similar to gastrulation in reptiles and birds (see Figure 8-11E). Gastrulation movements in the inner cell mass produce a primitive streak. Epiblast cells move medially through the primitive streak into the blastocoel, and individual cells then migrate laterally through the blastocoel to form mesoderm and endoderm. Endoderm cells (derived from the hypoblast) form a yolk sac devoid of yolk (since the mammalian embryos derive nutrients directly from the mother via the placenta).
Amphibians, reptiles, and birds, which have moderate to large amounts of yolk concentrated in the vegetal region of the egg, have evolved derived gastrulation patterns in which the yolk does not participate in gastrulation. Yolk is an impediment to gastrulation and consequently the gastrulation process occurs around (amphibians) or on top (reptiles and birds) of the vegetal yolk. Mammalian eggs are isolecithal, and thus one might expect them to have a gastrulation pattern similar to that of sea stars. Instead they have a pattern more suited to telolecithal eggs. The best explanation for this feature of mammalian egg development is common ancestry with birds and reptiles. Reptiles, birds, and mammals share a common ancestor whose eggs were telolecithal. Thus, all three groups inherited their gastrulation patterns from this common ancestor, and mammals subsequently evolved isolecithal eggs but retained the telolecithal pattern.
In Cnidaria and Ctenophora, only two germ layers are formed, endoderm and ectoderm. These animals are diploblastic. In all other metazoa, the mesoderm also appears, either from pouches of the archenteron or from other cells associated with endoderm formation. This three-layered condition is called triploblastic.
Support our developers




