Nuclear Equivalence
Mechanisms of
Development
Nuclear Equivalence
How does a developing embryo generate the multitude of many cell types of a complete multicellular organism from the starting point of a single diploid nucleus of a zygote? To many nineteenthcentury embryologists there seemed only one acceptable answer: as cell division ensued, hereditary material had to be parceled unequally to daughter cells. In this view, the genome gradually became broken into smaller and smaller units until finally only the information required to impart the characteristics of a single cell type remained. This became known as the Roux-Weismann hypothesis, after the two German embryologists who developed the concept.
However, in 1892 Hans Driesch discovered that if he mechanically shook apart a two-celled sea urchin into separate cells, both half-embryos developed into normal larvae. Driesch concluded that both cells contained all the genetic information of the original zygote. Still, this experiment did not settle the argument, because many embryologists believed that even if all cells contained complete genomes, the nuclei might become progressively modified in some way to dispense with the information they do not use in forming differentiated cells.
The efforts of Hans Driesch to disrupt egg development are poetically described by Peattie:“Behold Driesch grinding the eggs of Loeb’s favorite sea urchin up between plates of glass, pounding and breaking and deforming them in every way.And when he ceased from thus abusing them, they proceeded with their orderly and normal development. Is any machine conceivable, Driesch asks, which could thus be torn down . . . have its parts all disarranged and transposed, and still have them act normally? One cannot imagine it. But of the living egg, fertilized or not,we can say that there lie latent within it all the potentialities presumed by Aristotle, and all of the sculptor’s dream of form, yes, and the very power in the sculptor’s arm.”From Peattie, D. C. 1935. An Almanac for Moderns. New York,G. P. Putnams Sons.
Around the turn of the century
Hans Spemann introduced a new approach
to testing the Roux-Weismann
hypothesis. Spemann placed minute
ligatures of human hair around salamander
zygotes just as they were
about to divide, constricting them until
they were almost, but not quite, separated
into two halves (Figure 8-13).
The nucleus lay in one half of the partially
divided egg; the other side was
anucleate, containing only cytoplasm.
The egg then completed its first cleavage
division on the side containing the
nucleus; the anucleate side remained
undivided. Eventually, when the nucleated
side had divided into about 16
cells, one of the cleavage nuclei would
wander across the narrow cytoplasmic
bridge to the anucleate side. Immediately
this side began to divide.
With both halves of the embryo containing nuclei, Spemann drew the ligature tight, separating the two halves of the embryo. He then watched their development. Usually two complete embryos resulted (Figure 8-13A). Although the one embryo would have possessed only one-sixteenth the original nuclear material (according to the Roux-Weismann hypothesis), and the other contained fifteen-sixteenths, they both developed normally. The one-sixteenth embryo was initially smaller, but it caught up in size in about 140 days. This showed that a single nucleus selected from the 16-cell embryo contained a complete set of genes; all were equivalent.
Sometimes, however, Spemann observed that the nucleated half of the embryo developed only into an abnormal ball of “belly” tissue, although the half that received the delayed nucleus developed normally. Why should the more generously endowed fifteensixteenths embryo fail to develop and the small one-sixteenth embryo live? The explanation, Spemann discovered, depending on the position of the gray crescent, the pigment-free area that appears at the moment of fertilization. If one-half of the constricted embryo lacked a part of the gray crescent, it would not develop (Figure 8-13B).
Spemann’s delayed nucleation experiments served as compelling evidence for two important conclusions: (1) all cells contained the same nuclear information (thus disproving the Roux- Weismann hypothesis), and (2) cytoplasm in the area of the gray crescent must contain information essential for normal development.
If all nuclei are equivalent, what causes some cells to develop into neurons while others develop into skeletal muscle? In most animals (excluding insects), there are two major ways by which cells become committed to particular developmental fates: (1) cytoplasmic segregation of determinative molecules during cleavage and (2) interaction with neighboring cells (inductive interactions). All animals use both of these mechanisms to some extent to specify different cell types. However, in some animals cytoplasmic specification is dominant, whereas others rely predominantly on inductive interactions.
Nuclear Equivalence
How does a developing embryo generate the multitude of many cell types of a complete multicellular organism from the starting point of a single diploid nucleus of a zygote? To many nineteenthcentury embryologists there seemed only one acceptable answer: as cell division ensued, hereditary material had to be parceled unequally to daughter cells. In this view, the genome gradually became broken into smaller and smaller units until finally only the information required to impart the characteristics of a single cell type remained. This became known as the Roux-Weismann hypothesis, after the two German embryologists who developed the concept.
However, in 1892 Hans Driesch discovered that if he mechanically shook apart a two-celled sea urchin into separate cells, both half-embryos developed into normal larvae. Driesch concluded that both cells contained all the genetic information of the original zygote. Still, this experiment did not settle the argument, because many embryologists believed that even if all cells contained complete genomes, the nuclei might become progressively modified in some way to dispense with the information they do not use in forming differentiated cells.
The efforts of Hans Driesch to disrupt egg development are poetically described by Peattie:“Behold Driesch grinding the eggs of Loeb’s favorite sea urchin up between plates of glass, pounding and breaking and deforming them in every way.And when he ceased from thus abusing them, they proceeded with their orderly and normal development. Is any machine conceivable, Driesch asks, which could thus be torn down . . . have its parts all disarranged and transposed, and still have them act normally? One cannot imagine it. But of the living egg, fertilized or not,we can say that there lie latent within it all the potentialities presumed by Aristotle, and all of the sculptor’s dream of form, yes, and the very power in the sculptor’s arm.”From Peattie, D. C. 1935. An Almanac for Moderns. New York,G. P. Putnams Sons.
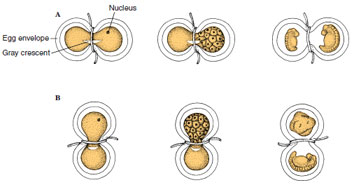 |
| Figure 8-13 Spemann’s delayed nucleation experiments. Two kinds of experiments were performed. A, Hair ligature was used to constrict an uncleaved fertilized newt egg. Both sides contained part of the gray crescent. The nucleated side alone cleaved until a descendant nucleus crossed over the cytoplasmic bridge. Then both sides completed cleavage and formed two complete embryos. B, Hair ligature was placed so that the nucleus and gray crescent were completely separated. The side lacking the gray crescent became an unorganized piece of belly tissue; the other side developed normally. |
With both halves of the embryo containing nuclei, Spemann drew the ligature tight, separating the two halves of the embryo. He then watched their development. Usually two complete embryos resulted (Figure 8-13A). Although the one embryo would have possessed only one-sixteenth the original nuclear material (according to the Roux-Weismann hypothesis), and the other contained fifteen-sixteenths, they both developed normally. The one-sixteenth embryo was initially smaller, but it caught up in size in about 140 days. This showed that a single nucleus selected from the 16-cell embryo contained a complete set of genes; all were equivalent.
Sometimes, however, Spemann observed that the nucleated half of the embryo developed only into an abnormal ball of “belly” tissue, although the half that received the delayed nucleus developed normally. Why should the more generously endowed fifteensixteenths embryo fail to develop and the small one-sixteenth embryo live? The explanation, Spemann discovered, depending on the position of the gray crescent, the pigment-free area that appears at the moment of fertilization. If one-half of the constricted embryo lacked a part of the gray crescent, it would not develop (Figure 8-13B).
Spemann’s delayed nucleation experiments served as compelling evidence for two important conclusions: (1) all cells contained the same nuclear information (thus disproving the Roux- Weismann hypothesis), and (2) cytoplasm in the area of the gray crescent must contain information essential for normal development.
If all nuclei are equivalent, what causes some cells to develop into neurons while others develop into skeletal muscle? In most animals (excluding insects), there are two major ways by which cells become committed to particular developmental fates: (1) cytoplasmic segregation of determinative molecules during cleavage and (2) interaction with neighboring cells (inductive interactions). All animals use both of these mechanisms to some extent to specify different cell types. However, in some animals cytoplasmic specification is dominant, whereas others rely predominantly on inductive interactions.
Support our developers




