How Amount and Distribution of Yolk Affect Cleavage
How Amount and Distribution
of Yolk Affect Cleavage
Eggs with very little yolk that is evenly
distributed in the egg are called isolecithal (Gr. isos, equal, + lekithos,
yolk). In such eggs, cleavage is holoblastic (Gr. holo, whole, + blastos,
germ), meaning that the cleavage
furrow extends completely through the
egg (see Figure 8-7A, C, and E).
Isolecithal eggs are found in a great
diversity of animals, including echinoderms,
tunicates, cephalochordates,
nemerteans, most molluscs, as well as
marsupial and placental mammals
(including humans).
Amphibian eggs (Figure 8-7B) are called mesolecithal (Gr. mesos, middle, + lekithos, yolk) because they have a moderate amount of yolk concentrated in the vegetal pole. The opposite animal pole contains mostly cytoplasm and very little yolk. Mesolecithal eggs also cleave holoblastically, but cleavage is substantially retarded in the yolk-rich vegetal pole. Each cleavage furrow begins at the animal pole and extends towards the vegetal pole. In axolotl salamanders, the cleavage furrow moves through the animal hemisphere at a rate of about 1 mm/min; it slows down to a rate of about 0.02 mm/min as it moves through the vegetal hemisphere. As a result, the second cleavage division begins at the animal pole while the first cleavage furrow is still slicing through the vegetal hemisphere. As cleavage progresses, the animal region becomes packed with numerous small cells, while the vegetal region contains relatively few, large, yolk-filled cells.
Eggs of birds, reptiles, most fishes, a few amphibians, cephalopod molluscs, and monotreme mammals are called telolecithal (Gr. telos, end, + lekithos, yolk) because they contain an abundance of yolk that is densely concentrated at the vegetal pole of the egg (refer to chick development in Figure 8-7D). The actively dividing cytoplasm is confined to a narrow discshaped mass lying on top of the yolk. Cleavage is partial, or meroblastic (Gr. meros, part, + blastos, germ), because the cleavage furrows cannot cut through the heavy yolk concentration, but instead stop at the border between the cytoplasm and yolk below.
Centrolecithal eggs, typical of insects and many other arthropods, also exhibit meroblastic cleavage (see Figure 8-8). These eggs have a large mass of centrally located yolk and cytoplasmic cleavage is limited to a surface layer of yolk-free cytoplasm while the yolkrich inner cytoplasm remains uncleaved.
Thus, yolk is an impediment to cleavage. In eggs with relatively little yolk (isolecithal and mesolecithal eggs), cleavage furrows can cut through the cytoplasm relatively easily and cleavages are therefore holoblastic. Once yolk becomes highly concentrated within portions of the egg (that is, telolecithal and centrolecithal eggs), cleavage furrows cannot penetrate the yolk and cytoplasmic cleavage is limited to relatively yolk-free areas, yielding a meroblastic type of cleavage.
How Amount of Yolk Affects Developmental Mode
The amount of yolk affects not only cleavage pattern, but also the developmental mode exhibited by embryos. Most animals receive no direct nourishment from the mother during embryonic development. However, the mother indirectly provides her eggs with nourishment by provisioning the egg (during oogenesis) with yolk, which fuels development until the offspring is able to obtain food on its own.
Zygotes of most aquatic invertebrates contain limited yolk for growth, and develop rapidly into a freeswimming, morphologically distinct larval stage, which is specialized to feed itself to sustain further development (see Figure 8-21). This is called indirect development because the larval stage is interposed in the developmental sequence between embryo and adult. The larva will later undergo a metamorphosis into the adult body form. Indirect development is also characteristic of most amphibians. Mammalian zygotes (such as those of the mouse, Figure 8-7E) contain little yolk but have evolved a strategy that allows them to bypass the larval stage. They develop a placental attachment to the mother through which they are nourished during the long gestation. This is an example of direct development. Another means of achieving direct development is seen in reptiles and birds, which have no larval stage or placental attachment but whose eggs are provisioned with enough yolk to support growth until hatching as juveniles (which generally resemble the adult in body form).
How Cleavage Is Affected by Different Inherited Patterns
Another important influence on a species’ pattern of cleavage is its inherited pattern of cell division. This effect is most apparent in isolecithal eggs, in which four major patterns of cleavage can be observed: radial holoblastic, spiral holoblastic, bilateral holoblastic, and rotational holoblastic cleavage. These different cleavage patterns are characteristic of different phylogenetic groups of animals.
In radial cleavage (so called because the embryonic cells are arranged in radial symmetry around the animalvegetal axis), each cleavage furrow is oriented either parallel or perpendicular to the animal-vegetal axis of the egg. In sea stars (Figure 8-7A), the first cleavage plane passes right through the animal vegetal axis, yielding two identical daughter cells (called blastomeres). For the second cleavage division, furrows form simultaneously in both blastomeres, and these also are oriented parallel to the animalvegetal axis (but perpendicular to the first cleavage furrow). Cleavage furrows next form simultaneously in the four daughter blastomeres, this time oriented perpendicular to the animalvegetal axis, yielding two tiers of four cells each. Subsequent cleavages yield an embryo composed of several tiers of cells. Radial cleavage also is seen in most amphibian embryos, although the pattern is altered a bit due to slowing of the cleavage furrow as it moves through the yolk.
Spiral cleavage (represented by
nemertean worm development in Figure
8-7C) is different from radial in two
important ways. Rather than dividing
parallel or perpendicular to the animalvegetal
axis, blastomeres cleave oblique
to this axis and typically produce quartets
of cells that come to lie, not on top
of each other, but in the furrows
between the cells. In addition, spirally
cleaving blastomeres pack themselves
tightly together much like a group of
soap bubbles, rather than just lightly
contacting each other as do many radially
cleaving blastomeres.
The importance of these two cleavage patterns extends well beyond the differences we have described. They are signals of a fundamental dichotomy, the early evolutionary divergence of bilateral metazoan animals into two separate lineages. Spiral cleavage is found in annelids, nemerteans, turbellarian flatworms, all molluscs except cephalopods, some brachiopods, and echiurans. These and several other invertebrate phyla are included in the Protostomia division of the animal kingdom. Radial cleavage is characteristic of the Deuterostomia division of the animal kingdom, a grouping that traditionally includes echinoderms (sea stars and their kin), hemichordates, and chordates. Other distinguishing developmental hallmarks of these two divisions are summarized in Figure 8-9.
Ascidians (also known as tunicates) are relatives of vertebrates, have isolecithal eggs, and exhibit a unique type of cleavage called bilateral cleavage. In ascidian eggs, the anteroposterior axis is defined prior to fertilization by the asymmetrical distribution of several cytoplasmic components (Figure 8-10). The first cleavage furrow passes through the animal-vegetal axis, dividing the asymmetrically distributed cytoplasm equally between the first two blastomeres. Thus, this first cleavage division separates the embryo into its future right and left sides, establishing its bilateral symmetry (hence the name bilateral holoblastic cleavage). Each successive division orients itself to this plane of symmetry, and the halfembryo formed on one side of the first cleavage is the mirror image of the half embryo on the other side.
Most mammals possess isolecithal eggs and a unique cleavage pattern called rotational cleavage, so called because of the orientation of blastomeres with respect to each other during the second cleavage division (see mouse development in Figure 8-7E). Cleavage in mammals is slower than in any other animal group. In humans, the first division is completed about 36 hours after fertilization (compared with about a minute and a half in sea urchins), and the next divisions follow at 12- to 24-hour intervals. As in most other animals, the first cleavage plane runs through the animal-vegetal axis to yield a two-cell embryo. However, during the second cleavage one of these blastomeres divides meridionally (that is, through the animal-vegetal axis) while the other divides equatorially (that is, perpendicular to the animal-vegetal axis). Thus, the cleavage plane in one blastomere is rotated 90 degrees with respect to the cleavage plane of the other blastomere (hence the name rotational cleavage). Furthermore, early divisions are asynchronous; all blastomeres do not divide at the same time. Thus, mammalian embryos may not increase regularly from two to four to eight blastomeres, but often contain odd numbers of cells. After the third division, the cells suddenly close into a tightly packed configuration, which is stabilized by tight junctions that form between outermost cells of the embryo. These outer cells form the trophoblast. The trophoblast is not part of the embryo proper but will form the embryonic portion of the placenta when the embryo implants in the uterine wall. Cells that actually give rise to the embryo proper form from the inner cells, called the inner cell mass (see blastula stage in Figure 8-11E).
Among animals that exhibit meroblastic cleavage, there are two major inherited patterns of cleavage. Telolecithal eggs of reptiles, birds, and most fish divide by discoidal cleavage. Because of the great mass of yolk in these eggs, cleavage is confined to a small disc of cytoplasm lying atop a mound of yolk (see chick development in Figure 8-7D). Early cleavage furrows carve this cytoplasmic disc to yield a single layer of cells called the blastoderm. Further cleavages divide the blastoderm into five to six layers of cells. By contrast, the centrolecithal eggs of insects undergo superficial cleavage (Figure 8-8). The centrally located mass of yolk restricts cleavage to the cytoplasmic rim of the egg. This pattern is highly unusual because cytoplasmic cleavage (cytokinesis) does not occur until after many rounds of nuclear division. After roughly eight rounds of mitosis in the absence of cytoplasmic division (yielding 256 nuclei), the nuclei migrate to the yolkfree periphery of the egg. A few of the nuclei at the posterior end of the egg become surrounded by cytoplasm to form the pole cells, which will give rise to germ cells of the adult. Next, the entire egg cell membrane folds inward, partitioning each nucleus into a single cell, and yielding a layer of cells at the periphery surrounding the mass of yolk (Figure 8-8). Thus, different groups of animals have evolved different mechanisms for dealing with large volumes of yolk. Because yolk is an impediment to cleavage, both these patterns avoid cleaving the yolk and instead confine cytoplasmic division to small regions of yolk-free cytoplasm.
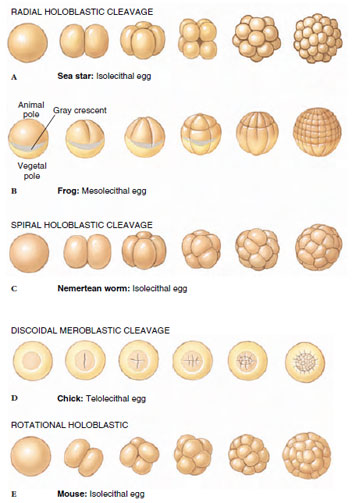 |
| Figure 8-7 Cleavage stages in sea star, frog, nemertean worm, chick, and mouse. |
Amphibian eggs (Figure 8-7B) are called mesolecithal (Gr. mesos, middle, + lekithos, yolk) because they have a moderate amount of yolk concentrated in the vegetal pole. The opposite animal pole contains mostly cytoplasm and very little yolk. Mesolecithal eggs also cleave holoblastically, but cleavage is substantially retarded in the yolk-rich vegetal pole. Each cleavage furrow begins at the animal pole and extends towards the vegetal pole. In axolotl salamanders, the cleavage furrow moves through the animal hemisphere at a rate of about 1 mm/min; it slows down to a rate of about 0.02 mm/min as it moves through the vegetal hemisphere. As a result, the second cleavage division begins at the animal pole while the first cleavage furrow is still slicing through the vegetal hemisphere. As cleavage progresses, the animal region becomes packed with numerous small cells, while the vegetal region contains relatively few, large, yolk-filled cells.
Eggs of birds, reptiles, most fishes, a few amphibians, cephalopod molluscs, and monotreme mammals are called telolecithal (Gr. telos, end, + lekithos, yolk) because they contain an abundance of yolk that is densely concentrated at the vegetal pole of the egg (refer to chick development in Figure 8-7D). The actively dividing cytoplasm is confined to a narrow discshaped mass lying on top of the yolk. Cleavage is partial, or meroblastic (Gr. meros, part, + blastos, germ), because the cleavage furrows cannot cut through the heavy yolk concentration, but instead stop at the border between the cytoplasm and yolk below.
Centrolecithal eggs, typical of insects and many other arthropods, also exhibit meroblastic cleavage (see Figure 8-8). These eggs have a large mass of centrally located yolk and cytoplasmic cleavage is limited to a surface layer of yolk-free cytoplasm while the yolkrich inner cytoplasm remains uncleaved.
Thus, yolk is an impediment to cleavage. In eggs with relatively little yolk (isolecithal and mesolecithal eggs), cleavage furrows can cut through the cytoplasm relatively easily and cleavages are therefore holoblastic. Once yolk becomes highly concentrated within portions of the egg (that is, telolecithal and centrolecithal eggs), cleavage furrows cannot penetrate the yolk and cytoplasmic cleavage is limited to relatively yolk-free areas, yielding a meroblastic type of cleavage.
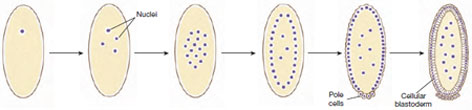 |
| Figure 8-8 Superficial cleavage in a Drosophila embryo. The zygote nucleus at first divides repeatedly in the yolk-rich endoplasm by mitosis without cytokinesis. After several rounds of mitosis, most nuclei migrate to the surface where they are separated by cytokinesis into separate cells. Some nuclei migrate to the posterior pole to form the primordial germ cells, called pole cells. Several nuclei remain in the endoplasm where they will regulate breakdown of yolk products. The cellular blastoderm stage corresponds to the blastula stage of other embryos. |
How Amount of Yolk Affects Developmental Mode
The amount of yolk affects not only cleavage pattern, but also the developmental mode exhibited by embryos. Most animals receive no direct nourishment from the mother during embryonic development. However, the mother indirectly provides her eggs with nourishment by provisioning the egg (during oogenesis) with yolk, which fuels development until the offspring is able to obtain food on its own.
Zygotes of most aquatic invertebrates contain limited yolk for growth, and develop rapidly into a freeswimming, morphologically distinct larval stage, which is specialized to feed itself to sustain further development (see Figure 8-21). This is called indirect development because the larval stage is interposed in the developmental sequence between embryo and adult. The larva will later undergo a metamorphosis into the adult body form. Indirect development is also characteristic of most amphibians. Mammalian zygotes (such as those of the mouse, Figure 8-7E) contain little yolk but have evolved a strategy that allows them to bypass the larval stage. They develop a placental attachment to the mother through which they are nourished during the long gestation. This is an example of direct development. Another means of achieving direct development is seen in reptiles and birds, which have no larval stage or placental attachment but whose eggs are provisioned with enough yolk to support growth until hatching as juveniles (which generally resemble the adult in body form).
How Cleavage Is Affected by Different Inherited Patterns
Another important influence on a species’ pattern of cleavage is its inherited pattern of cell division. This effect is most apparent in isolecithal eggs, in which four major patterns of cleavage can be observed: radial holoblastic, spiral holoblastic, bilateral holoblastic, and rotational holoblastic cleavage. These different cleavage patterns are characteristic of different phylogenetic groups of animals.
In radial cleavage (so called because the embryonic cells are arranged in radial symmetry around the animalvegetal axis), each cleavage furrow is oriented either parallel or perpendicular to the animal-vegetal axis of the egg. In sea stars (Figure 8-7A), the first cleavage plane passes right through the animal vegetal axis, yielding two identical daughter cells (called blastomeres). For the second cleavage division, furrows form simultaneously in both blastomeres, and these also are oriented parallel to the animalvegetal axis (but perpendicular to the first cleavage furrow). Cleavage furrows next form simultaneously in the four daughter blastomeres, this time oriented perpendicular to the animalvegetal axis, yielding two tiers of four cells each. Subsequent cleavages yield an embryo composed of several tiers of cells. Radial cleavage also is seen in most amphibian embryos, although the pattern is altered a bit due to slowing of the cleavage furrow as it moves through the yolk.
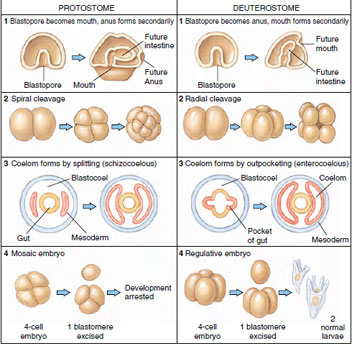 |
| Figure 8-9 Developmental tendencies of protostomes and deuterostomes. These tendencies are much modified in some groups, for example, the vertebrates. Cleavage in mammals is rotational rather than radial; in reptiles, birds, and many fishes cleavage is discoidal. Vertebrates have also evolved a derived form of coelom formation that is basically schizocoelous. |
The importance of these two cleavage patterns extends well beyond the differences we have described. They are signals of a fundamental dichotomy, the early evolutionary divergence of bilateral metazoan animals into two separate lineages. Spiral cleavage is found in annelids, nemerteans, turbellarian flatworms, all molluscs except cephalopods, some brachiopods, and echiurans. These and several other invertebrate phyla are included in the Protostomia division of the animal kingdom. Radial cleavage is characteristic of the Deuterostomia division of the animal kingdom, a grouping that traditionally includes echinoderms (sea stars and their kin), hemichordates, and chordates. Other distinguishing developmental hallmarks of these two divisions are summarized in Figure 8-9.
Ascidians (also known as tunicates) are relatives of vertebrates, have isolecithal eggs, and exhibit a unique type of cleavage called bilateral cleavage. In ascidian eggs, the anteroposterior axis is defined prior to fertilization by the asymmetrical distribution of several cytoplasmic components (Figure 8-10). The first cleavage furrow passes through the animal-vegetal axis, dividing the asymmetrically distributed cytoplasm equally between the first two blastomeres. Thus, this first cleavage division separates the embryo into its future right and left sides, establishing its bilateral symmetry (hence the name bilateral holoblastic cleavage). Each successive division orients itself to this plane of symmetry, and the halfembryo formed on one side of the first cleavage is the mirror image of the half embryo on the other side.
 |
| Figure 8-10 Bilateral cleavage in ascidian embryos. The first cleavage division divides the asymmetrically distributed cytoplasm evenly between the first two blastomeres, establishing the future right and left sides of the adult animal. Bilateral symmetry of the embryo is maintained through subsequent cleavage divisions. |
Most mammals possess isolecithal eggs and a unique cleavage pattern called rotational cleavage, so called because of the orientation of blastomeres with respect to each other during the second cleavage division (see mouse development in Figure 8-7E). Cleavage in mammals is slower than in any other animal group. In humans, the first division is completed about 36 hours after fertilization (compared with about a minute and a half in sea urchins), and the next divisions follow at 12- to 24-hour intervals. As in most other animals, the first cleavage plane runs through the animal-vegetal axis to yield a two-cell embryo. However, during the second cleavage one of these blastomeres divides meridionally (that is, through the animal-vegetal axis) while the other divides equatorially (that is, perpendicular to the animal-vegetal axis). Thus, the cleavage plane in one blastomere is rotated 90 degrees with respect to the cleavage plane of the other blastomere (hence the name rotational cleavage). Furthermore, early divisions are asynchronous; all blastomeres do not divide at the same time. Thus, mammalian embryos may not increase regularly from two to four to eight blastomeres, but often contain odd numbers of cells. After the third division, the cells suddenly close into a tightly packed configuration, which is stabilized by tight junctions that form between outermost cells of the embryo. These outer cells form the trophoblast. The trophoblast is not part of the embryo proper but will form the embryonic portion of the placenta when the embryo implants in the uterine wall. Cells that actually give rise to the embryo proper form from the inner cells, called the inner cell mass (see blastula stage in Figure 8-11E).
Among animals that exhibit meroblastic cleavage, there are two major inherited patterns of cleavage. Telolecithal eggs of reptiles, birds, and most fish divide by discoidal cleavage. Because of the great mass of yolk in these eggs, cleavage is confined to a small disc of cytoplasm lying atop a mound of yolk (see chick development in Figure 8-7D). Early cleavage furrows carve this cytoplasmic disc to yield a single layer of cells called the blastoderm. Further cleavages divide the blastoderm into five to six layers of cells. By contrast, the centrolecithal eggs of insects undergo superficial cleavage (Figure 8-8). The centrally located mass of yolk restricts cleavage to the cytoplasmic rim of the egg. This pattern is highly unusual because cytoplasmic cleavage (cytokinesis) does not occur until after many rounds of nuclear division. After roughly eight rounds of mitosis in the absence of cytoplasmic division (yielding 256 nuclei), the nuclei migrate to the yolkfree periphery of the egg. A few of the nuclei at the posterior end of the egg become surrounded by cytoplasm to form the pole cells, which will give rise to germ cells of the adult. Next, the entire egg cell membrane folds inward, partitioning each nucleus into a single cell, and yielding a layer of cells at the periphery surrounding the mass of yolk (Figure 8-8). Thus, different groups of animals have evolved different mechanisms for dealing with large volumes of yolk. Because yolk is an impediment to cleavage, both these patterns avoid cleaving the yolk and instead confine cytoplasmic division to small regions of yolk-free cytoplasm.
Support our developers




