Gradualism
Gradualism
Darwin’s theory of gradualism opposed arguments for the sudden origin of species. Small differences, resembling those that we observe among organisms within populations today, are the raw material from which the different major forms of life evolved. This theory shares with Lyell’s uniformitarianism the notion that we must not explain past changes by invoking unusual catastrophic events that are not observed today. If new species originated in single, catastrophic events, we should be able to see such events happening today and we do not. Instead, what we observe in natural populations are small, continuous changes in phenotypes. Such continuous changes can produce major differences among species only by accumulating over many thousands to millions of years. A simple statement of Darwin’s theory of gradualism is that accumulation of quantitative changes leads to qualitative change.
Mayr (see Figure 6-18) makes an important distinction between populational gradualism and phenotypic gradualism. Populational gradualism states that new traits become established in a population by increasing their frequency initially from a small fraction of the population to a majority of the population. Populational gradualism is well established and is not controversial. Phenotypic gradualism states that new traits, even those that are strikingly different from ancestral ones, are produced in a series of small, incremental steps.
Phenotypic Gradualism
Phenotypic gradualism was controversial when Darwin first proposed it, and it is still controversial. Not all phenotypic changes are small, incremental ones. Some mutations that appear during artificial breeding change the phenotype substantially in a single mutational step. Such mutations traditionally are called “sports.” Sports that produce dwarfing are observed in many species, including humans, dogs, and sheep, and have been used by animal breeders to achieve desired results; for example, a sport that deforms the limbs was used to produce ancon sheep, which cannot jump hedges and are therefore easily contained (Figure 6-23). Many colleagues of Darwin who accepted his other theories considered phenotypic gradualism too extreme. If sporting mutations can be used in animal breeding, why must we exclude them from our evolutionary theory? In favor of gradualism, some have replied that sporting mutations always have negative side-effects that would prevent them from surviving in natural populations. Indeed, it is questionable whether ancon sheep, despite their attractiveness to farmers, would propagate successfully in the presence of their long-legged relatives without human intervention.
Punctuated equilibrium
When we view Darwinian gradualism on a geological timescale, we may expect to find in the fossil record a long series of intermediate forms connecting the phenotypes of ancestral and descendant populations (Figure 6-24). This predicted pattern is called phyletic gradualism. Darwin recognized that phyletic gradualism is not often revealed by the fossil record. Studies conducted since Darwin’s time generally have not revealed the continuous series of fossils predicted by phyletic gradualism. Is the theory of gradualism therefore refuted by the fossil record? Darwin and others claim that it is not, because the fossil record is too imperfect to preserve transitional series. Although evolution is a slow process by our standards, it is rapid relative to the rate at which good fossil deposits accumulate. Others have argued, however, that abrupt origins and extinctions of species in the fossil record force us to conclude that phyletic gradualism is rare.
Niles Eldredge and Stephen Jay
Gould proposed punctuated equilibrium
to explain the discontinuous
evolutionary changes observed throughout
geological time. Punctuated equilibrium
states that phenotypic evolution
is concentrated in relatively brief
events of branching speciation, followed
by much longer intervals of
evolutionary stasis (Figure 6-25). Speciation
is an episodic event, having a
duration of approximately 10,000 to
100,000 years. Because species may
survive for 5 million to 10 million
years, the speciation event is a “geological
instant,” representing 1% or less
of a species’ life span. Ten thousand
years is plenty of time, however, for
Darwinian evolution to accomplish
dramatic changes. A small fraction of
the evolutionary history of a group
therefore contributes most of the morphological
evolutionary change that
we observe.
The process of allopatric speciation by founder events provides a possible explanation for punctuated equilibria. Remember that founder-induced speciation requires the breaking of genetic equilibrium in a small, geographically isolated population. Such small populations have very little chance of being preserved in the fossil record. After a new genetic equilibrium forms and stabilizes, the new population may increase in size, thereby increasing the likelihood that some of its members will be preserved as fossils. Founder-induced speciation cannot be the exclusive cause of punctuated equilibrium, however, because punctuated equilibrium may be observed in groups where speciation by founder events is unlikely.
Evolutionists who lamented the imperfect state of the fossil record were treated in 1981 to the opening of an uncensored page of fossil history in Africa. Peter Williamson, a British paleontologist working in fossil beds 400 m deep near Lake Turkana, documented a remarkably clear record of speciation in freshwater snails. The geology of the Lake Turkana basin reveals a history of instability. Earthquakes, volcanic eruptions, and climatic changes caused the waters to rise and fall periodically, sometimes by hundreds of feet. Thirteen lineages of snails show long periods of stability interrupted by relatively brief periods of rapid change in shell shape when snail populations were fragmented by receding waters. These populations diverged to produce new species that then remained unchanged through thick deposits before becoming extinct and being replaced by descendant species. The transitions occurred within 5000 to 50,000 years. In the few meters of sediment where speciation occurred, transitional forms were visible. Williamson’s study conforms well to the punctuated equilibrium model of Eldredge and Gould.
 |
| Figure 6-23 The ancon breed of sheep arose from a “sporting mutation” that a used dwarfing of legs. Many of his contemporaries criticized Darwin for his claim that such mutations are not important in the process of evolution by natural selection. |
Darwin’s theory of gradualism opposed arguments for the sudden origin of species. Small differences, resembling those that we observe among organisms within populations today, are the raw material from which the different major forms of life evolved. This theory shares with Lyell’s uniformitarianism the notion that we must not explain past changes by invoking unusual catastrophic events that are not observed today. If new species originated in single, catastrophic events, we should be able to see such events happening today and we do not. Instead, what we observe in natural populations are small, continuous changes in phenotypes. Such continuous changes can produce major differences among species only by accumulating over many thousands to millions of years. A simple statement of Darwin’s theory of gradualism is that accumulation of quantitative changes leads to qualitative change.
Mayr (see Figure 6-18) makes an important distinction between populational gradualism and phenotypic gradualism. Populational gradualism states that new traits become established in a population by increasing their frequency initially from a small fraction of the population to a majority of the population. Populational gradualism is well established and is not controversial. Phenotypic gradualism states that new traits, even those that are strikingly different from ancestral ones, are produced in a series of small, incremental steps.
Phenotypic Gradualism
Phenotypic gradualism was controversial when Darwin first proposed it, and it is still controversial. Not all phenotypic changes are small, incremental ones. Some mutations that appear during artificial breeding change the phenotype substantially in a single mutational step. Such mutations traditionally are called “sports.” Sports that produce dwarfing are observed in many species, including humans, dogs, and sheep, and have been used by animal breeders to achieve desired results; for example, a sport that deforms the limbs was used to produce ancon sheep, which cannot jump hedges and are therefore easily contained (Figure 6-23). Many colleagues of Darwin who accepted his other theories considered phenotypic gradualism too extreme. If sporting mutations can be used in animal breeding, why must we exclude them from our evolutionary theory? In favor of gradualism, some have replied that sporting mutations always have negative side-effects that would prevent them from surviving in natural populations. Indeed, it is questionable whether ancon sheep, despite their attractiveness to farmers, would propagate successfully in the presence of their long-legged relatives without human intervention.
Punctuated equilibrium
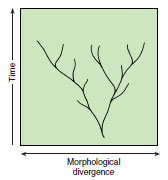 |
| Figure 6-24 The gradualist model of evolutionary change in morphology, viewed as proceeding more or less steadily through geological time (millions of years). Bifurcations followed by gradual divergence led to speciation. |
When we view Darwinian gradualism on a geological timescale, we may expect to find in the fossil record a long series of intermediate forms connecting the phenotypes of ancestral and descendant populations (Figure 6-24). This predicted pattern is called phyletic gradualism. Darwin recognized that phyletic gradualism is not often revealed by the fossil record. Studies conducted since Darwin’s time generally have not revealed the continuous series of fossils predicted by phyletic gradualism. Is the theory of gradualism therefore refuted by the fossil record? Darwin and others claim that it is not, because the fossil record is too imperfect to preserve transitional series. Although evolution is a slow process by our standards, it is rapid relative to the rate at which good fossil deposits accumulate. Others have argued, however, that abrupt origins and extinctions of species in the fossil record force us to conclude that phyletic gradualism is rare.
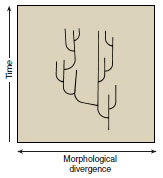 |
| Figure 6-25 The punctuated equilibrium model sees evolutionary change being concentrated in relatively rapid bursts of branching speciation (lateral lines) followed by prolonged periods of no change throughout geological time (millions of years). |
The process of allopatric speciation by founder events provides a possible explanation for punctuated equilibria. Remember that founder-induced speciation requires the breaking of genetic equilibrium in a small, geographically isolated population. Such small populations have very little chance of being preserved in the fossil record. After a new genetic equilibrium forms and stabilizes, the new population may increase in size, thereby increasing the likelihood that some of its members will be preserved as fossils. Founder-induced speciation cannot be the exclusive cause of punctuated equilibrium, however, because punctuated equilibrium may be observed in groups where speciation by founder events is unlikely.
Evolutionists who lamented the imperfect state of the fossil record were treated in 1981 to the opening of an uncensored page of fossil history in Africa. Peter Williamson, a British paleontologist working in fossil beds 400 m deep near Lake Turkana, documented a remarkably clear record of speciation in freshwater snails. The geology of the Lake Turkana basin reveals a history of instability. Earthquakes, volcanic eruptions, and climatic changes caused the waters to rise and fall periodically, sometimes by hundreds of feet. Thirteen lineages of snails show long periods of stability interrupted by relatively brief periods of rapid change in shell shape when snail populations were fragmented by receding waters. These populations diverged to produce new species that then remained unchanged through thick deposits before becoming extinct and being replaced by descendant species. The transitions occurred within 5000 to 50,000 years. In the few meters of sediment where speciation occurred, transitional forms were visible. Williamson’s study conforms well to the punctuated equilibrium model of Eldredge and Gould.
Support our developers




