Homology and Phylogenetic Reconstruction
Homology and Phylogenetic
Reconstruction
Darwin recognized the major source of evidence for common descent in the concept of homology. Darwin’s contemporary, Richard Owen (1804 to 1892), used this term to denote “the same organ in different organisms under every variety of form and function.” A classic example of homology is the limb skeleton of vertebrates. Bones of vertebrate limbs maintain characteristic structures and patterns of connection despite diverse modifications for different functions (Figure 6- 13). According to Darwin’s theory of common descent, the structures that we call homologies represent characteristics inherited with some modification from a corresponding feature in a common ancestor.
Darwin devoted an entire book, The Descent of Man and Selection in
Relation to Sex, largely to the idea that
humans share common descent with
apes and other animals. This idea was
repugnant to the Victorian world,
which responded with predictable outrage
(Figure 6-14). Darwin built his
case mostly on anatomical comparisons
revealing homology between
humans and apes. To Darwin, the
close resemblances between apes and
humans could be explained only by
common descent.
Throughout the history of all forms of life, evolutionary processes generate new characteristics that are then inherited by subsequent generations. Every time a new feature arises on an evolving lineage, we see the origin of a new homology. That homology gets transmitted to all descendant lineages unless it is subsequently lost. The pattern formed by the sharing of homologies among species provides evidence for common descent and allows us to reconstruct the branching evolutionary history of life. We can illustrate such evidence using a phylogenetic tree for a group of large, ground-dwelling birds (Figure 6-15). A new skeletal homology arises on each of the lineages shown (descriptions of specific homologies are not included because they are highly technical). The different groups of species located at the tips of the branches contain different combinations of these homologies, which reflect ancestry. For example, ostriches show homologies 1 through 5 and 8, whereas kiwis show homologies 1, 2, 13, and 15. Branches of the tree combine these species into a nested hierarchy of groups within groups (see Classification and Phylogeny of Animals). Smaller groups (species grouped near terminal branches) are contained within larger ones (species grouped by basal branches, including the trunk of the tree). If we erase the tree structure but retain patterns of homology observed in the living species, we are able to reconstruct the branching structure of the entire tree. Evolutionists test the theory of common descent by observing the patterns of homology present within all groups of organisms. The pattern formed by all homologies taken together should specify a single branching tree that represents the evolutionary genealogy of all living organisms.
The nested hierarchical structure
of homology is so pervasive in the living
world that it forms the basis for our
systematic classification of all forms of
life (genera grouped into families,
families grouped into orders, and
other categories). Hierarchical classification
even preceded Darwin’s theory
because this pattern is so evident,
but it was not explained adequately
before Darwin. Once the idea of common
descent was accepted, biologists
began investigating the structural, molecular,
and chromosomal homologies
of animal groups. Taken together, the
nested hierarchical patterns uncovered
by these studies have permitted us to
reconstruct evolutionary trees of many
groups and to continue investigating
others. Use of Darwin’s theory of common
descent to reconstruct the evolutionary
history of life and to classify
animals is the subject of Classification and Phylogeny of Animals.
Note that the earlier evolutionary hypothesis that life arose many times, forming unbranched lineages, predicts linear sequences of evolutionary change with no nested hierarchy of homologies among species. Because we do observe nested hierarchies of homologies, that hypothesis is rejected. Note also that because the creationist argument is not a scientific hypothesis, it can make no testable predictions about any pattern of homology.
Darwin recognized the major source of evidence for common descent in the concept of homology. Darwin’s contemporary, Richard Owen (1804 to 1892), used this term to denote “the same organ in different organisms under every variety of form and function.” A classic example of homology is the limb skeleton of vertebrates. Bones of vertebrate limbs maintain characteristic structures and patterns of connection despite diverse modifications for different functions (Figure 6- 13). According to Darwin’s theory of common descent, the structures that we call homologies represent characteristics inherited with some modification from a corresponding feature in a common ancestor.
 |
| Figure 6-14 This 1873 advertisement for Merchant’s Gargling Oil ridicules Darwin’s theory of the common descent of humans and apes, which received only limited acceptance by the general public during Darwin’s lifetime. |
Throughout the history of all forms of life, evolutionary processes generate new characteristics that are then inherited by subsequent generations. Every time a new feature arises on an evolving lineage, we see the origin of a new homology. That homology gets transmitted to all descendant lineages unless it is subsequently lost. The pattern formed by the sharing of homologies among species provides evidence for common descent and allows us to reconstruct the branching evolutionary history of life. We can illustrate such evidence using a phylogenetic tree for a group of large, ground-dwelling birds (Figure 6-15). A new skeletal homology arises on each of the lineages shown (descriptions of specific homologies are not included because they are highly technical). The different groups of species located at the tips of the branches contain different combinations of these homologies, which reflect ancestry. For example, ostriches show homologies 1 through 5 and 8, whereas kiwis show homologies 1, 2, 13, and 15. Branches of the tree combine these species into a nested hierarchy of groups within groups (see Classification and Phylogeny of Animals). Smaller groups (species grouped near terminal branches) are contained within larger ones (species grouped by basal branches, including the trunk of the tree). If we erase the tree structure but retain patterns of homology observed in the living species, we are able to reconstruct the branching structure of the entire tree. Evolutionists test the theory of common descent by observing the patterns of homology present within all groups of organisms. The pattern formed by all homologies taken together should specify a single branching tree that represents the evolutionary genealogy of all living organisms.
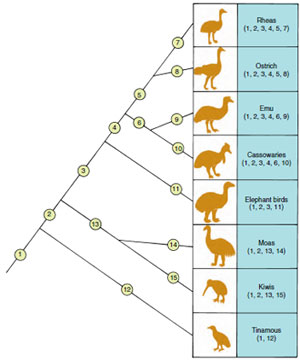 |
| Figure 6-15 The phylogenetic pattern specified by twelve homologous structures in the skeletons of a group of flightless birds. Homologous features are numbered 1 through 12 and are marked both on the branches of the tree on which they arose and on the birds that have them. If you were to erase the tree structure, you would be able to reconstruct it without error from the distributions of homologous features shown for the birds at the terminal branches. |
Note that the earlier evolutionary hypothesis that life arose many times, forming unbranched lineages, predicts linear sequences of evolutionary change with no nested hierarchy of homologies among species. Because we do observe nested hierarchies of homologies, that hypothesis is rejected. Note also that because the creationist argument is not a scientific hypothesis, it can make no testable predictions about any pattern of homology.
Support our developers




