Speciation and Extinction through Geological Time
Speciation and Extinction
through Geological Time
Evolutionary change at the second tier provides a new perspective on Darwin’s theory of natural selection. A species has two possible evolutionary fates: it may give rise to new species or become extinct without leaving descendants. Rates of speciation and extinction vary among lineages, and lineages that have the highest speciation rates and lowest extinction rates produce the greatest diversity of living forms. The characteristics of a species may make it more or less likely than others to undergo speciation or extinction events. Because many characteristics are passed from ancestral to descendant species (analogous to heredity at the organismal level), lineages whose characteristics increase the probability of speciation and confer resistance to extinction should come to dominate the living world. This species-level process that produces differential rates of speciation and extinction among lineages is analogous in many ways to natural selection. It represents an expansion of Darwin’s theory of natural selection.
Species selection is the differential survival and multiplication of species through geological time based on variation among lineages in emergent, species-level properties. These species-level properties include mating rituals, social structuring, migration patterns, geographic distribution, and all other properties that emerge at the species level. Descendant species usually resemble their ancestors in these properties. For example, a “harem” system of mating in which a single male and several females compose a breeding unit characterizes some mammalian lineages but not others. We expect speciation rates to be enhanced by social systems that promote founding of new populations by small numbers of individuals. Certain social systems may increase the likelihood that a species will survive environmental challenges through cooperative action. Such properties would be favored by species selection over geological time.
Effect macroevolution is similar
to species selection except that differential
speciation and extinction among
lineages is caused by variation in
organismal-level properties (such as
specialized versus generalized feeding)
rather than species-level properties. Organisms that specialize in
eating a restricted range of foods, for
example, may be subjected more readily
than generalized feeders to geographic
isolation among populations,
because areas where their preferred
food is scarce or absent will function
as geographic barriers to dispersal.
Such geographic isolation could generate
more frequent opportunities for
speciation to occur throughout geological
time. The fossil records of two
major groups of African antelopes
demonstrate this result (Figure 6-33). A
lineage of specialized grazers that contains
blesboks, hartebeests, and wildebeests
shows high speciation and
extinction rates; since the late Miocene,
33 extinct and 7 living species are
found, representing at least 18 events of
branching speciation and 12 terminal
extinctions. In contrast, a lineage of generalist
grazers and browsers that contains
impalas shows neither branching
speciation nor terminal extinction during
this same interval of time. Interestingly,
although these two lineages differ
greatly in speciation rates, extinction
rates, and species diversity, they do not
differ significantly in total number of
individual animals alive today.
Evolutionary change at the second tier provides a new perspective on Darwin’s theory of natural selection. A species has two possible evolutionary fates: it may give rise to new species or become extinct without leaving descendants. Rates of speciation and extinction vary among lineages, and lineages that have the highest speciation rates and lowest extinction rates produce the greatest diversity of living forms. The characteristics of a species may make it more or less likely than others to undergo speciation or extinction events. Because many characteristics are passed from ancestral to descendant species (analogous to heredity at the organismal level), lineages whose characteristics increase the probability of speciation and confer resistance to extinction should come to dominate the living world. This species-level process that produces differential rates of speciation and extinction among lineages is analogous in many ways to natural selection. It represents an expansion of Darwin’s theory of natural selection.
Species selection is the differential survival and multiplication of species through geological time based on variation among lineages in emergent, species-level properties. These species-level properties include mating rituals, social structuring, migration patterns, geographic distribution, and all other properties that emerge at the species level. Descendant species usually resemble their ancestors in these properties. For example, a “harem” system of mating in which a single male and several females compose a breeding unit characterizes some mammalian lineages but not others. We expect speciation rates to be enhanced by social systems that promote founding of new populations by small numbers of individuals. Certain social systems may increase the likelihood that a species will survive environmental challenges through cooperative action. Such properties would be favored by species selection over geological time.
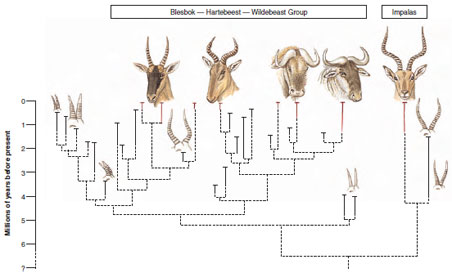 |
| Figure 6-33 Contrasting diversity between two major groups of African antelopes. Higher speciation and extinction rates in the group containing the blesboks, hartebeests, and wildebeests is attributed to greater specialization in feeding relative to the impalas, an example of effect macroevolution. |
Support our developers




